Не виходили наші пращури з Африки

Знайшли древні скелетні залишки, але як дізнатися, це «вперше», або завтра знайдуть ще древніші? Наскільки надійні датування, які насправді зовсім не надійні і практично завжди оскаржуються? Є десятки антропологічних ознак, які так чи інакше приміряють на поняття Homo sapiens і на поняття «анатомічно сучасна людина», але теорія це одне (хоча загальновизнаною повної класифікації все одно немає), а на практиці ці ознаки застосувати повною мірою практично неможливо - зазвичай знаходять лише фрагменти скелета, часто без лицьових кісток, і для найбільш древніх кісткових залишків практично завжди спостерігаються якісь «архаїчні» ознаки.
І далі в справу вступає те, що називають сумлінністю науковця. Ставки великі - кожен новий скелет або його фрагмент, який дає можливість проголосити його «найдавнішим з відомих» Homo sapiens або АСЧ, стає всесвітньою сенсацією, з усіма відповідними наслідками у вигляді наукових нагород, великих фінансових грантів, виборами в престижні академії наук. Тому, на жаль, так нерідкі спотворення даних, що описуються в академічній та іншій пресі, не кажучи про популярні видання, такі ласі на сенсації. Датування у наукових виданнях деколи завищуються, архаїчні ознаки «замазуються», і стає зовсім важко розібратися, де справжні дані, а де фантазії авторів. Потрібні перехресні дослідження, які рідкісні. Нарешті, чимало просто некваліфікованих робіт, особливо в галузі популяційної генетики, або робіт, орієнтованих на наперед заданий результат.
Про це й буде наше оповідання. А саме, про те, як важко пробити стіну ангажованих досліджень, які «заточені» на нібито вихід «анатомічно сучасної людини» нібито з Африки, і що насправді показують дослідження, часто тих же авторів, але своєрідно ними інтерпретуються. Стіна до того ж цементується ідеологічними міркуваннями, по яких треба будь-якою ціною показати «африканське походження людини», а хто знаходить інші дані і проводить інші інтерпретації, той «расист». Стіна зміцнюється і тим, що практично всі статті ангажованих дослідників, а це більшість популяційних генетиків, починаються фразою «Як відомо, анатомічно сучасна людина вийшла з Африки». Тобто установка йде з самого початку. Це різко підвищує ймовірність публікації статті в академічному виданні.
Ось кілька прикладів, взятих з назв академічних статей, або з перших фраз вступів в статті:
Походження людини: вихід з Африки (назва статті; Tattersal, 2009);
Еволюція людини і вихід з Африки (з назви статті; Stewart and Stringer, 2012);
Африканське походження чоловічого (генетичного) різноманітності (з назви статті; Cruciani et al, 2011);
Африканське походження сучасних людей Східної Азії (з назви статті; Ke et al, 2001);
... анатомічно сучасні люди прибули до Європи з Африки щонайменше 45 тисяч років тому, слідом за поширенням людей з Африки (Moorjani et al, 2011);
Вважають, що сучасні люди походять в східній Африці (Henn et al, 2011);
Загальновизнано, що анатомічно сучасні люди походять в Африці (Hammer et al, 2011);
Африка, прабатьківщина всіх сучасних людей (Lachance et al, 2012);
... розходження анатомічно сучасних людей з Африки було приблизно 44 тисячі років тому (Underhill et al, 2000);
Сучасні люди відбулися в Африці приблизно 200 тисяч років тому (Campbell and Tishkoff, 2010);
... Анатомічно сучасні люди походять з невеликої ізольованої популяції в Африці 150-200 тисяч років тому (Patin et al, 2009);
Суб-Сахара і північно-східна Африка - найбільш ймовірні регіони походження людини і коридор в інший світ (Arredi et al, 2004);
... розходження людини почалося в Африці (Ramachandran et al., 2005).
Нижче в цій роботі буде показано, що всі ці положення, і аналогічні їм, які відображаються в десятках і сотнях академічних та інших статей - невірні.
Питання - як наука про походження людини дійшла до такого життя? Як міг скластися «консенсус фахівців», заснований на однобокій і накатаній інтепретаціі під заздалегідь задану відповідь? Як взагалі в науці могла скластися ситуація, коли інші обгрунтовані інтерпретації тих же або інших даних зустрічаються вираженою агресією, політичними звинуваченнями, неприкритими негативними емоціями? Чому «вихід з Африки» став релігією, заснованої на вірі, яка доказів не потребує?
До 1980-х років обговорення африканського походження людини йшло мляво, і були по суті маргінальними. Розглядати це серйозно заважали дві обставини. Перше - було визнано, що далеким пращуром сучасної людини був Homo erectus, людина прямоходяча, який з'явилася кілька мільйонів років тому, можливо, в Африці, але було відомо, що вже майже два мільйони років тому він поширився по всій Євразії. Тому Homo sapiens, людина розумна, міг стати його нащадком де завгодно. Друге - було показано, що найближчий родич АСЧ, неандерталець, в Африці не жив. Тому загальний пращур сучасної людини і неандертальця, який жив за різними даними між 600 і 300 тисяч років тому, виходить, що в Африці теж не жив. До того ж у неандертальця була світла шкіра, і на цьому ми зупинимося нижче. Тому африканське походження сучасної людини вимагає прибуття світлошкірого безпосереднього пращура людини в Африку, скажімо, 500-300 тисяч років тому, далі було самостійне, еволюційне придбання їм чорної шкіри, інакше в Африці не вижити, а потім вихід його з Африки і зворотне самостійне перетворення чорної шкіри у світлу. На цей рахунок була навіть придумана дотепна гіпотеза про роль вітаміну D в самостійному (без схрещування зі світлошкірими людьми, яких бути за межами Африки не могло, інакше концепція руйнується) перетворення чорношкірих в світлошкірих, але ця гіпотеза ніколи не була підтверджена експериментально. Так і залишилася умоглядною.
Так що там у статті Ребекки Канн 1987? Що лягло в основу нової релігії? З чого все це почалося? Давайте подивимося.
Основоположна стаття Канн та ін ( 1987 ) про «вихід з Африки»
У вступі до статті - ні слова про Африку і нібито зародження там людства. Тобто стаття позиціонує себе як перший в цьому відношенні. Експериментальна частина статті - визначення нуклеотидної послідовності мтДНК з 147 жінок з п'яти основних регіонів:
Африка - 20 осіб (двоє народжені до півдня від Сахари, решта - чорношкірі мешканці США, зазвичай метиси з домішкою Y-ДНК чоловіків - європеоїдів, але у цих 18 людях «передбачається африканська мтДНК, на що також вказує характер мутацій фрагментів мтДНК»);
Азія (Китай, В'єтнам, Лаос, Філіппіни, Індонезія, Полінезія/Тонга ) - 34 людини;
Європеоїди (Європа, Північна Африка, Близький Схід) - 46 осіб;
Австралійські аборигени - 21 людина;
Нова Гвінея - 26 осіб.
Всі мтДНК були розщеплені на фрагменти за допомогою ферментів - рестриктаз, отримані в сумі 467 незалежних ділянок мтДНК, з яких 195 мали відмінності хоча б у однієї людини з усіх 147. Іншими словами, було ідентифіковано 195 поліморфних ділянок мтДНК. У середньому аналіз проводили на 9% від усієї мтДНК. Загалом, для того часу, 25 років тому це була досить просунута в технічному відношенні робота.
Далі провели попарні зіставлення отриманих фрагментів ДНК між усіма 147 учасницями, і знайшли, що ці попарні відмінності варіюються від нуля до 1.3 мутацій на кожні 100 нуклеотидів (0 до 1.3% відмінностей), із загальною усередненою величиною 0.32% відмінностей. Але треба було показати, що ці відмінності найбільше у африканців, тому всі п'ять популяцій розділили на кластери, виходячи з груп попарних відмінностей у кожній популяції. Вийшло, що
46 європейських мтДНК розходяться на 36 кластерів,
34 азіатських мтДНК розходяться на 27 кластерів,
21 австралійських мтДНК розходяться на 15 кластерів,
26 мтДНК з Нової Гвінеї розходяться на 7 кластерів,
а 20 африканських мтДНК визначили в один кластер, ухваливши, що раз людство вийшло з Африки, то там повинен бути всього один кластер. Так і записали в примітці до таблиці в статті, де у всіх вказано багато кластерів, а у африканців всього один.
Якщо хто розуміє, що зробили автори статті, то бажання читати далі вже пропадає. Бо далі автори стали визначати, яка популяція старше інших, для того на кластери і розбивали. Але фокус був у тому, що вік кожної популяції розраховували з різноманітності між кластерами всередині кожної популяції, а у африканців він автоматично був заданий найбільший, тому що у них кластер всього один. У всіх різноманітність гасилось в кожному кластері, а у африканців нічого не гасилось, кластер - один. Так і вийшло, ось «розмаїття» по кластерам:
Африка: 0.36%
Азія: 0.21%
Австралія: 0.17%
Нова Гвінея: 0.11%
Європа: 0.09%
Далі автори перевели ці «розмаїття» в хронологічні показники, а саме в роки, коли ці території були вперше заселені. Для цього взяли такі цифри для калібрування: заселення Австралії сталося 40 тисяч років тому, заселення Нової Гвінеї 30 тисяч років тому, заселення Америки 12 тисяч років тому, і отримали, що мутації в мтДНК виникають із середньою швидкістю 2-4% (тобто 2 -4 мутації на кожні 100 нуклеотидів) на мільйон років. Звідси автори статті вирахували середній «вік» кластерів в популяції :
Африка: 90-180 тисяч років
Азія: 53-105
Австралія: 43-85
Нова Гвінея: 28-55
Європа: 23-45
Зробили незграбно, але цифри вийшли досить розумні (в межах 100%-вої похибки). Як показали подальші дослідження вже інших авторів, у тому числі й мої, про які буде розказано нижче, африканські ДНК - лінії пішли починаючи приблизно з 160 тисяч років тому, плюс кілька архаїчних африканських ліній (гаплогрупи А0 і А00) віком приблизно 180 і 210 тисяч років, відповідно; азіатські і європейські лінії - починаючи з 64 тисяч років тому, Австралія - приблизно з 45-50 тисяч років тому, а найдавніші кісткові залишки сучасної людини в Європі датуються 45 тисяч років тому (Benazzi et al, 2011; Higham et al, 2011). Зрозуміло, що автори провели розрахунки з точністю плюс-мінус 100%, але проте загальна картина схоплена більш менш вірно.
Подібним же чином автори розрахували, що загальний пращурк всіх мтДНК жив 143-285 тисяч років тому, а оскільки загальний пращур всіх африканських мтДНК жив, за їх розрахунками, 90-180 тисяч років тому, тобто древніше всіх (хоча і перетинається за віком у межах похибки розрахунків), стало бути, тоді ж він і вийшов з Африки.
Чи помічаєте підміну понять? Автори розраховують, що люди поза Африкою походять від більш недавнього загального пращура, і постулюють, що, значить, він вийшов з Африки. У підсумку, узгоджують автори і це ж пишуть у Абстракті, жінка, загальний пращур всіх мтДНК на планеті, як «постулюється» (!), жила 200 тисяч років тому (це вже трансформація величини 143-285 тисяч років тому), і «ймовірно» (!) вона жила в Африці.
Ось з цієї статті все і почалося. Повторюю, що я не збагну, як така стаття змогла пройти рецензентів і бути опублікована в журналі Nature, з цими «постулюється» і «ймовірно», і за відсутності будь-яких даних про вихід сучасного людства з Африки, але саме так ця стаття і стала сприйматися і засобами інформації, і в популяційної генетики, а звідти і в науці і серед обивателя - що, значить, неспростовно доведено, що сучасна людина вийшла з Африки. Інших генетичних доказів фактично не було, та й навіщо? Все ж вже доведено, чи не так?
Принципові помилки творців і прихильників
концепції «вихід людства з Африки»
Є основна помилка, яка постійно переслідує популяційних генетиків. Якщо одна популяція генетично «різноманітніша» іншої, тобто в сукупності древніша, вони, як правило, вважають, що вона - пращурна по відношенню до другої. Але це зовсім не так. Тут треба дивитися сукупність факторів, а не перестрибувати до висновку. Наприклад, старший брат «різноманітніше» молодшого, але це зовсім не означає, що молодший - нащадок старшого. Просто у них є спільний пращур, їх батько. Це ж відноситься до різноманітних генеалогічних побудов, і якщо ми почнемо зіставляти племінників і відсувати їх спільних пращурів до загального дідуся, прадідуся, пра-прадідуся і так далі, то побачимо, що гілки нащадків можуть відходити від загального генеалогічного дерева в різні часи, але не можна їх порівнювати «за віком» лінійно, безпосередньо один з одним, потрібно неодмінно знати, коли жив їхній загальний пращур.
Це ясно, якщо подивитися на звичайне дерево. Поруч сидять товста гілка і молода гілочка, але зовсім не обов'язково, що молода виходить зі старої. Найчастіше вони незалежні до стовбура, їх загальний пращур - стовбур, або гілка ще більшої товщини. Концепція загального пращура в статті 1987 взагалі не зачіпалася. Типова помилка попгенетиків - це «що бачу, те й співаю». Раз зараз живуть в Африці - значить, завжди там жили. Те, що загальний пращур африканців і неафріканців міг жити поза Африки, і в давні часи туди мігрував - ними навіть і не розглядається.
Є ще одна принципова помилка підходу попгенетиків, заснованної на порівнянні «розмаїття». Різноманітність інформативно в даному сенсі, як каже термодинаміка, тільки в замкнутих системах. Нью-Йорк значно «різноманітніше», ніж, скажімо, Бостон, але чи означає це, що Бостон - нащадок Нью-Йорка, вийшов з нього? Москва «різноманітніше» Новгорода, але нащадок чи Новгород Москви? Зовсім ні. Швидше, навпаки. Різноманітність виходить найчастіше від змішування різних популяцій, тому що система відкрита. Ось в Нью-Йорку і в Москві намішано, і накопичилася велика «різноманітність». Африка - теж відкрита система. Туди в давнину і у відносно недавній час просунулися багато міграцій самих різних гаплогрупп, от і «різноманітність». Навіть гаплогруппа R1b у своїй частині просунулася близько 5 тисяч років тому, тепер живуть в Камеруні і Чаді (Cruciani et al, 2010), чорношкірі, тому що змішалися з місцевими красунями. А гаплогруппа так і залишилася, R1b. Додали вони в африканську «різноманітність»? Безумовно, як і безліч подібних міграцій в Африку. Час від часу з'являються академічні статті, в яких описується «вхід в Африку». Остання стаття - в тому ж журналі Nature в серпні 2013 (Hayden, 2013), в якій описані тільки що виявлені міграції популяцій в Африку 3000 років тому і 900-1800 років тому. Додали вони до «різноманітності»? Безумовно. Тим більше що пішли на південь від Сахари, звідки і брали проби мтДНК автори статті 1987 року.
Таку ж помилку робить Аткінсон у недавній статті (Atkinson, 2011), в якій він пише: «генетичне і фенотипічне різноманітність зменшується з віддаленням від Африки... що підтримує гіпотезу про африканське походження людини». Подивимося на діаграму, яка буде пояснена нижче. Зліва - африканська гілка, праворуч - неафриканська. Розмаїтість (тобто старовину) зменшується зліва направо, але зовсім не тому, що ліва гілка - пращурна. Вони обидві виходять із загального пращура, який, як буде показано нижче, в Африці не жив.
Ще ілюстрація з падінням різноманітності з віддаленням від Африки. Вік гаплогрупп А в Африці - приблизно 160 тисяч років, після розбіжності від альфа-гаплогрупи. Вік гаплогрупп R1a і R1b на видаленні від Африки - 20 тис років і 16 тис років, відповідно, вони утворилися в Центральній Азії (Klyosov and Rozhanskii, 2012a; Klyosov, 2012). Різноманітність падає від Африки до Центральної Азії? Падає. Чи тому, що R1a і R1b походять від африканських гаплогрупп А? Абсолютно ні. Це - незв'язані події і системи.
Аналогія - якщо в одній частині міста знаходиться будинок престарілих, то «різноманітність» у ньому найвища. Чи означає це, що все в місті включаючи дитячий сад навпроти, походять від будинку престарілих? Зовсім не обов'язково. Це - незв'язані події і системи. Це могло б бути вірно, якби система була замкнутою, тобто в місто ніхто не в'їжджав протягом сотень років. Але в реальності в місто в'їжджають тисячі людей, які не мають ніякого відношення до тих, хто в будинку престарілих і хто в дитячому садку, так і в перший привозять престарілих з усієї країни та з-за кордону. Хоча якщо виміряти - різноманітність у ньому найвище. А ось пращурної немає.
Ось приклади «лінійного мислення» щодо «розмаїття»:
... Різноманітність гаплотипів найвище в Африці (Hellenthal et al., 2008);
Африка має не тільки найвищий рівень генетичної варіації в світі, але також має значну різноманітність в мовах, культурі, природних умовах (Campbell and Tishkoff, 2010).
Все це так, але до головної тези про походження людства нібито в Африці відношення не має. Це - всього пара прикладів із сотень такого ж роду.
Для обговорення «генетичної різноманітності» треба знати генезис популяцій, їх історію, а не просто формально це «різноманітність» вимірювати у різних популяцій і «лінійно» зіставляти. Це, повторюю, бич популяційних генетиків. Чому це? Слабка наукова школа, іншого пояснення немає.
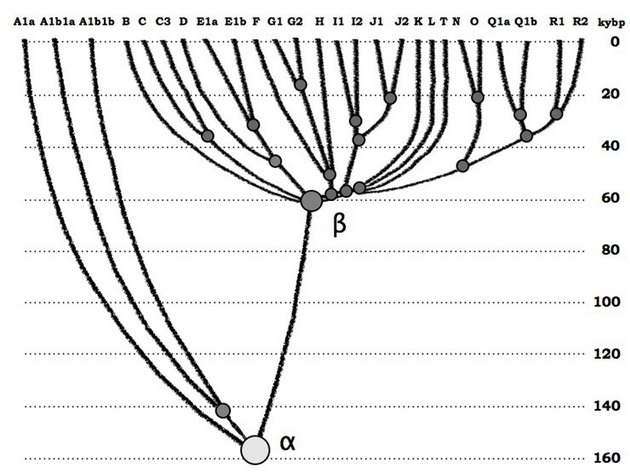
Діаграма еволюції гаплогрупп сучасного людства. На горизонтальній осі - основні гаплогрупи Y-хромосоми людства, на вертикальній - абсолютна шкала часу. Спільний пращур альфа-гаплогрупи жив приблизно 160 тисяч років тому, загальний пращур бета-гаплогрупи (або гаплогрупп від В до Т) - 64±6 тисяч років тому (зі статті AA Klyosov and IL Rozhanskii, Advances in Anthropology, 2012b). На діаграмі не показані архаїчні африканські лінії А00 і А0 (остання зараз замінила в номенклатурі лінії A1b на діаграмі ліворуч), оновлене дерево гаплогрупп буде показано нижче.
Є ще (як мінімум) одна причина нижчого «розмаїття» у неафріканців. Приблизно 64 тисячі років тому їх пращури пройшли «пляшкове горлечко популяції». Інакше кажучи, в результаті якогось катаклізму неафріканці майже всі загинули, або виродилися, і вижила їх тільки невелика група. Образно кажучи, вижили в кінцевому підсумку нащадки всього лише однієї пари, і ось до них сходяться всі генеалогічні лінії практично всіх чоловіків на планеті. Що за катаклізм або інша напасть, як, наприклад, мор, трапилася - невідомо, і найбільшу вагу мають дві гіпотези - виверження вулкана Тоба, найбільше з відомих в історії людства, приблизно 70 тисяч років тому, і похолодання в північній гемісфері. Кліматологи стверджують, що ранг катастрофічності похолодання вище, ніж ранг виверження Тоби. Як там не було, ось що вийшло (див. наведену діаграму).
Зрозуміло, що якщо вимірювати «різноманітність» гілок зліва (африканської за нинішнім проживанням її представників) і справа (неафриканських, теж за нинішнім проживання), то перша буде древнє. Але права гілка не виходить з лівої, у них - загальний пращур, альфа-гаплогруппа. Як видно, діаграма пояснює всі результати статті Канн 1987 года, але без виходу з Африки. Інші докази правильності цієї діаграми будуть приведені нижче.
І пішли десятки і сотні академічних статей, перша фраза в яких зазвичай була - «Як відомо, анатомічно сучасна людина вийшла з Африки приблизно 70 тисяч років тому». Втім, і це датування було «плаваючим», і нижче даються приклади різних датувань «виходу з Африки» у різних статтях. Невеликий секрет - жодна з них не була дійсно розрахунковою. Всі вони - просто так, «зі стелі». Та й не було до останнього часу відповідного розрахункового апарату, а той, що був - читач уже побачив, який він і яка там точність.
50 тисяч років тому (Thomson et al, 2000);
50-60 тисяч років тому (Shi et al., 2010);
50-60 тисяч років тому (Mellars, 2011);
50-70 тисяч років тому (Hudjasov et al., 2007);
50-70 тисяч років тому (Stoneking & Delfin, 2010);
60 тисяч років тому (Li & Durbin, 2011);
60 тисяч років тому (Henn et al., 2011);
60 тисяч років тому (Wei et al., 2013);
60-70 тисяч років тому (Ottoni et al., 2010);
60-80 тисяч років тому (Forster, 2004);
54 ± 8 тисяч років тому (Forster et al., 2001);
60 тисяч років тому (Stewart & Stringer, 2012);
45-50 тисяч років тому (Fernandes et al., 2012);
50-65 тисяч років тому (Behar et al., 2008);
50-60 тисяч років тому (Cann, 2013);
60 тисяч років тому (Chiaroni et al., 2009);
50-75 тисяч років тому (Patin et al., 2009);
50 тисяч років тому (Edmonds et al., 2004);
45 тисяч років тому (Moorjani et al., 2011);
50-70 тисяч років тому (Xue et al., 2005);
70-80 тисяч років тому (Majumder, 2010);
40 тисяч років тому (Campbell and Tishkoff, 2010);
50 тисяч років тому (Poznik et al, 2013);
55-70 тисяч років тому (Soares et al., 2009);
між 40 і 70 тисяч років тому (Sahoo et al., 2006);
між 35 і 89 тисяч років тому (Underhill et al., 2000);
між 80 і 50 тисяч років тому (Yotova et al., 2011);
між 50 і 100 тисяч років тому (Hublin, 2011);
між 27-53 і 58-112 тисяч років тому (Carrigan and Hammer, 2006);
70-60 тисяч років тому (Curnoe et al., 2012);
~ 110 тисяч років тому (Francalacci et al, 2013);
200 тисяч років тому (Hayden, 2013).
Насправді ніякої дати обгрунтовано привести не можна. Її просто немає. Та й всі ці датування вище нікому не потрібні, вони нічого не дають і не відповідають по суті ні на яке питання. Це - все та ж мантра.
Що кажуть насправді експериментальні дані
і їх більш широка інтерпретація?
Зупинимося на час з критикою, і подивимося - а що ж є? Якщо сучасні неафріканці - НЕ нащадки давніх африканців, то звідки це випливає? Чиї вони нащадки?
Археологічні, палеонтологічні дані по Африці доведеться з жалем відкинути. Вони пізнавальні із загальних міркувань, але ми не знаємо, чи були у знайдених кісткових залишків взагалі виживші нащадки. Можливо, ми спостерігаємо залишки терміновавших ліній. Поки не будуть встановлені гаплогрупи і гаплотипи тих кісткових залишків, вони нам нічого не скажуть про спадкоємність цих еволюційних ліній. Далі, ми не знаємо, звідки ці кісткові залишки там з'явилися. Можливо, їх близькі пращури мігрували в Африку. Дійсно, якщо можливо було вийти з Африки, то так само можливо було туди і увійти. Більше того, відомі багато прикладів міграцій в Африку. Багато датуваннь древніх кісткових залишків невірні, і приклади будуть дані нижче. Багато заявлені як «стародавні Homo sapiens» мають виражені архаїчні ознаки, та їх віднесення до Homo sapiens взагалі є спірним або просто невірним. Багато знахідок відносяться взагалі не до кісткових залишків, а до стоянок, печер, знайдених там раковин, кам'яним знаряддям праці. Невідомо, хто там взагалі був, і знайдена там охра теж ні про що не говорить. Неандертальці Євразії теж застосовували кам'яні знаряддя і охру для своїх цілей.
Так що до розглянутого питання про те, як співвідносяться стародавні африканці та неафріканці, більше підходить розглядати ДНК тих і інших. Якщо ці дані підтримуються і археологією-антропологією - це чудово, але поки таких даних мало, якщо вони взагалі є. Подивимося на них.
Дані по ДНК можна розглядати в трьох варіантах, які в принципі зобов'язані давати взаємно узгоджені дані. Це (1) гаплотипи і гаплогрупи Y-хромосоми людини, (2) мтДНК людини, і (3) геном людини. Останнє фактично означає картину необоротних мутацій в ДНК, яка піддається інтерпретації щодо еволюції людини, показуючи напрям перетікання мутацій і появи нових у ході еволюційного розвитку. Наприклад, в геномі як неандертальця, так і сучасної людини є багато одних і тих же мутацій, які є і в ДНК шимпанзе. Це означає, що дані мутації - від загального пращура людини і шимпанзе. Але якщо в нас є і мутації від неандертальця, яких немає у шимпанзе - то це може означати, що неандерталець - наш прямий пращур. Такі мутації або не встановлені, або їх дуже небагато і вони спірні. Дані зараз переглядаються. Нещодавно проголошені 1-4% як нібито у сучасної людини від неандертальця теж зараз переглядаються. Швидше за все, вони невірні.
Таким же чином як у неафріканцях, так і в африканцях є спільні мутації від загального пращура з шимпанзе. Їх багато, і вони нецікаві при вирішенні питання про те, чи походимо ми від африканців. Ці мутації повинні бути відфільтровані. А от чи є в нас такі мутації, які є у африканців, але немає у шимпанзе - ось це питання, на яке має бути відповідь. Ця відповідь, якщо така буде отриманою, повинна узгоджуватися з даними за гаплотипом і гапллогруппам Y-хромосоми і по мтДНК. Ось так має будуватися дослідження того питання, які еволюційні шляхи сучасного людства.
Насправді, ця відповідь вже отримана - немає в нас «африканських» мутацій, придбаних ними за останні 150-200 тисяч років. Мутації від загального пращура з шимпанзе, яким мільйони років, в наших ДНК є, і багато, а мутацій від африканців, придбаних ними за останні 160 тисяч років, в наших ДНК немає.
Про це і піде мова.
Отже, що говорять на цей рахунок експериментальні дані? Почнемо з гаплотипів і гаплогрупп людини, що проектуються на часи більше 100 тисяч років тому. Проектованих - тому що копалин гаплотипів і гаплогрупп того часу немає. Ще не виявлені, завдання технічно дуже складнае, тому що за такі часи ДНК людини розкладається майже повністю, особливо під впливом мікроорганізмів. ДНК неандертальця (точніше, неандерталки) давністю 45 тисяч років виділена і в значній мірі розшифрована, а ось ДНК людини 160 тисяч років тому - це завдання на порядок є важчим.
Як чинять у таких випадках? Визначають гаплотипи у сучасних популяцій чоловіків, якщо аналіз проводять по Y-хромосомі. Але вибірку аналізують не по якійсь «новогвінейської» або «африканської» популяції, яка може бути зовсім різнорідною, а серед носіїв певного субклада гаплогрупи, тобто серед людей, об'єднаних конкретним набором мутацій. Вони - родичі, і для них досить точно розраховується, коли жив їхній загальний пращур. Наприклад, серед сучасних африканців є досить представницька група людей, яка за класифікацією потрапляє в субклад A1b1b2b, позначений кольором (третій знизу) на дереві субкладів гаплогрупи А нижче. Це дерево показує ієрархію субкладів гаплогрупи A, тобто еволюцію гаплогрупи А. Видно, як галузиться дерево - від стовбура в сторону відійшла найдавніша гаплогруппа А00, її гілки (субклади) поки невідомі. Стовбур продовжує гаплогруппа А0-Т, яка розійшлася на два субклада - А0 і А1; А1 в свою чергу розійшлася на А1А і A1b; A1b - на A1b1 і ВТ Зведена гаплогруппа ВТ, як буде ще показазано, дуже віддалена від гаплогрупп серії «А», та й у серії «А» незрозуміло, які субклади можуть бути названі африканськими за походженням. Поки складається, що тільки гаплогрупи А00 і А0, то є перша і третя зверху (відійшли убік від стовбурової гілки, що веде від загального пращура з шимпанзе мільйони років тому, до неафриканських гаплогруп ВТ) на показаному нижче дереві, і субклади останньої (A0a, A0b, A0a1, A0a2, A0a1a і A01ab) можуть розглядатися як африканські за походженням або по прибуттю в Африку більше 100 тисяч років тому. Решта, починаючи з А1 (що знаходиться на тій же стовбуровій гілці), роздвоюється на африканські (відходять в сторону) і імовірно неафриканські (стовбурові) гілки.

Давайте розглянемо ще раз, більш чітко, як галузиться дерево гаплогрупп, як кожна гілка розходиться на вілки, і як одна частина вілки йде (мігрує) в Африку, а інша частина залишається поза Африкою, і знову розходиться на чергову вілку. Інакше кажучи, в Африку міграції приходили хвилями. У підсумку простежується неафриканський стовбур, провідний до нас з вами, читачу, і від якого у бік відходять африканські пагони. Ми від цих пагонів не відбулися. Тут треба зауважити, що терміни «ствол», «стовбурова» і «відходять в сторону» обрані умовно, і можна, навпаки, назвати африканські гілки стовбуровими, а неафриканські - відходять в сторону. Ці поняття насправді симетричні.
Вілка 1 - від основного еволюційного Y-хромосомного «ствола», що йде від спільних пращурів з приматами (шимпанзе, горила, орангутанг, макака) приблизно 300-600 тисяч років тому відходить гілка неандертальців (Homo neanderthalensis); вони не були африканцями, у всякому разі, їх слідів в Африці не виявлено, тому можна вважати, що і загальний стовбур 300-600 тисяч років тому був неафриканським родом Homo.
Вілка 2 - від стовбура приблизно 210 тисяч років тому відходить найбільш древня з поки виявлених гілок букета гаплогрупп А, гаплогруппа A00 (зараз всі її виявлені носії живуть в Африці у складі племені Mbo, або є афро-американцями; ніяких відомостей про їх антропології або анатомії виявити не вдалося, в статті, яка призводить їх гаплотипи, про це ні слова).
Вілка 3 - стовбур досягає гаплогрупи А0 -Т (імовірно неафриканської), яка розходиться приблизно 180 тисяч років тому на африканську гаплогрупи А0 і імовірно неафриканську А1; іншими словами, від стовбура відходить чергова африканська гаплогруппа А0 .
Вілка 4 - неафриканська гаплогруппа А1 розходиться на африканську А1А і імовірно неафриканську А1b; іншими словами, від стовбура відходить чергова африканська гаплогруппа А1А.
Вілка 5 - неафриканська гаплогруппа A1b розходиться на африканську А1b1 і неафриканську BT (бета-гаплогруппа на першій діаграмі); іншими словами, від стовбура відходить чергова африканська гаплогруппа А1b1.
Тепер - дуже важливе положення нашого розгляду. Вілки 3, 4 і 5 розходяться від гаплогрупп А0-Т, А1 і А1b, відповідно.
Від першої в сторону відходить А0 (яка знайдена в Африці) і А1 (носії якої поки ніде не знайдені). Ми, неафріканці, походимо від А1 (і не походимо від А0, її мутацій в нас немає).
Від А1 в бік відходить А1А (яка знайдена в Африці) і A1b (носії якої поки ніде не знайдені). Ми, неафріканці, походимо від неї. У нашій Y-хромосомі - мутації від A1b, але не від A1a.
Від A1b в сторону відходить A1b1 (яка знайдена в Африці, Європі та Азії) і BT, з якої вийшли всі неафриканські гаплогрупи, в тому числі основні європейські гаплогрупи R1a, R1b, I1, I2, N1c1.
Тому в описі вілок вище я скрізь пишу «імовірно неафриканська гаплогруппа» A0-T, A1, A1b.
Таким чином, де б не жили першопращури неандертальців і ті, з ким вони розійшлися в ході еволюції (тобто ті, які продовжували «основний стовбур» еволюційного дерева Y-хромосоми), носії гаплогрупп А00, А0, A1a, A1b1 мігрували від них в Африку, і продовжували свою еволюцію там, приймаючи численних пізніших мігрантів в Африку і тим самим збільшуючи африканську «різноманітність».
В цілому, можна нарахувати чотири основних древніх міграцій в Африку протягом останніх кількох сот тисяч років - гаплогруппа А00 приблизно 210 тисяч років тому, гаплогруппа А0 приблизно 180 тисяч років тому, гаплогруппа А1А приблизно 160 тисяч років тому, гаплогруппа A1b1 приблизно 70 тисяч років тому. Зрозуміло, відбувалися і більш пізні міграції, наприклад 3000 і 900-1800 років тому, описані в роботі (Hayden, 2013), які теж збільшували «генетичну різноманітність» в Африці, так що «різноманітність» - не аргумент «прабатьківщини».
Я вже згадав, що носії гаплогрупи A1b1 живуть і в Африці, і в Європі, і в Азії. Мабуть, тому субклад A1b1b2b - M13 в Проекті гаплогрупи А виявляється найбільш численним. Він розходиться на дві основні гілки - арабську і європейську. Хто був родоначальником цих гілок і де він жив - ми не знаємо, але гілка досить неглибока, тобто вона відносно недавно пройшла пляшкове горлечко популяції. Її гаплотипи являють собою цінне джерело інформації, оскільки поміщають навіть неглибоку (за часом) гілку в поле всіх гаплотипів людства. Гаплотипи і після пляшкового горлечка популяції не могли самозародитися, вони могли тільки продовжити еволюцію від найдавніших спільних пращурів. Аналіз гаплотипів з використанням самих «повільних», найстабільніших 22 маркерів Y-хромосоми (Klyosov, 2011), показує, що загальний пращур арабської гілки субклада A1b1b2b мав гаплотип
12 11 11 9 11 10 10 9 12 12 7 12 8 0 13 11 16 9 14 9 11 11
а загальний пращур європейської гілки мав гаплотип
12 11 11 9 11 10 10 9 12 12 7 10 8 0 13 11 16 10 14 9 11 11
Між ними всього три мутації, що поміщає загального пращура арабської та європейської гілок приблизно на 7170 років тому, з похибкою плюс-мінус п'ять відсотків. Для цілей нашого опису ці розрахунки поки не дуже важливі, оскільки видно, що наведені вище гаплотипи близькі один до одного.
Порівняймо ці гаплотипи з пращурним африканським гаплотипом групи A00:
13 11 12 10 11 16 10 9 14 14 8 8 8 9 12 11 12 8 12 12 11 11
Це зіставлення показує різницю вже в 30 і 29 мутацій, тобто розводить спільних пращурів цих гаплотипів як мінімум на 286-308 тисяч років (формули розрахунків опубліковані в роботі [Klyosov, 2011, 2012]), і поміщає загального пращура гаплогрупи А00 приблизно на 210 тисяч років тому. Залежність між числом мутацій і згодом не лінійна, а статична, оскільки на великих часах частина мутацій повертається назад, і на це при розрахунках вводиться відповідна статистична поправка (Klyosov, 2009; Klyosov, 2012). Гаплотипи гаплогрупи А00 отримані у чорношкірого племені Мбо, що живе в Камеруні, і у афро-американця, імовірно вивезеного століття тому з того ж племені (Mendez et al, 2013).
Якщо ми тепер порівняємо ці гаплотипи з пращурним гаплотипом гаплогрупи В
11 12 11 11 11 10 11 8 16 16 8 10 8 12 10 11 15 8 12 11 12 11
то ми побачимо 29 мутацій від гаплогрупи А00, і практично стільки ж - 29 і 27 мутацій - від арабської та європейської гілок гаплогрупи A1b1b2b. Це - як мінімум 286-248 років між загальними пращурами гаплогрупп А і В. Цей колосальний поділ у часі ніяк не дозволяє гаплогруппі В бути нащадком гаплогрупи А. А ось мати загального пращура 160 тисяч років тому і розійтися від нього на 250-300 тисяч років - можна. Це знову узгоджується з діаграмою вище. Не можна ці гаплогрупи зіставляти «лінійно» тільки тому, що вони знаходяться візуально поруч на дереві гаплотипів, як не можна зіставляти гілки дерева в лісі «лінійно» , за відстанню між ними, тільки тому, що вони опинилися поруч. А поруч можуть виявитися гілки берези і ялини, що росте по сусідству.
Отже, гаплогруппа В дуже віддалена від гаплогрупи А, на 27-29-30 мутацій. А от від європейських (значною мірою) гаплогрупп R1a і R1b вона віддалена не так сильно, всього на 12 і 10 мутацій, відповідно:
11 12 13 11 11 12 11 9 15 16 8 10 8 12 10 12 12 8 12 11 11 12 (R1b - M269)
12 12 11 11 11 11 11 8 17 17 8 10 8 12 10 12 12 8 12 11 11 12 (R1a - Z280)
Самі ж ці гаплотипи (R1b і R1a) розділені всього 8 мутаціями, що відповідає часу життя їх спільного пращура (гаплогруппа R1) приблизно 26 тисяч років тому. Спільний пращур гаплогрупи В жив близько 50 тисяч років тому, і він не утворився з гаплогрупи А, вони - незалежні ДНК - генеалогічні лінії, що виходять з одного спільного пращура - альфа-гаплогрупи, 160 тисяч років тому.
У Європі живуть і інші носії гаплогрупи А, правда, їх поки знайдено небагато. Кілька років тому вийшла наукова стаття під назвою «Африканці в Йоркширі?» (King et al, 2007), в якій описано сімейство носіїв гаплогрупи А в Англії, які й не підозрювали, що у них африканські коріння по чоловічій лінії. Базовий 17 - маркерний гаплотип у них виявився наступний (у порядку маркерів DYS393, 390, 1 , 391, 388, 439, 389-1, 392, 389-2, 437, 438, 434, 435, 436, 460, 461, 462):
14 23 17 10 10 11 12 11 17 14 8 12 12 11 11 12 12
А у арабської гілки субклада A1b1b2b, описаного вище
13 21 15 9 11 12 13 11 18 16 10 9 11 11 13 11 13
Між ними - 20 мутацій на 17 маркерах, що відповідає як мінімум 19 тисячам років до їх загального пращу, англійських та арабських гаплотипів гаплогрупи А. Хто в цьому випадку куди пересувався - або в Африку, або з Африки - сказати неможливо. Сценарії могли бути будь-які. Прихильник ж концепції «вихід з Африки» тут же скаже, що вони вийшли з Африки. Ось така психологічна установка.
Дебати останніх двох років щодо наведеної діаграми
Коли стаття, що містить представлену вище діаграму та її інтепретації, була опублікована в травні 2012 року в журналі «Advances in Anthropology» (Успіхи антропології), це спочатку викликало неприйняття у популяційних генетиків. Конкретно, неприйняття викликали три основні висновки: (1) африканські та неафриканські ДНК-лінії розійшлися приблизно 160 тисяч років тому, і між ними є відповідна значна відстань; (2) неафриканські ДНК-лінії не є нащадками африканських гаплогрупп А00, А0, А з субкладамі; і, як наслідок, (3) ніякого «виходу з Африки» у сучасного людства не було, в усякому разі в останні 200 тисяч років. А якщо й було, то було туди-сюди, зустрічними міграціями, і сучасне людство чергова «туди-сюди» міграція не породила. У всякому разі, в даному відношенні вони еквівалентні.
Треба сказати, що це неприйняття в академічній науковій пресі взагалі не висловлювалося. Популяційні генетики як води в рот набрали. Пристрасті кипіли на англомовних форумах, у неформальних дискусіях. Було оголошено, що ця діаграма і, відповідно, її висновки повністю суперечать консенсусу про вихід людства з Африки, і суперечать всім опублікованим діаграммам і деревам гаплогрупп, отриманим за допомогою геномних досліджень. Було також оголошено, що неафриканські лінії виходять з гаплогрупп з індексом «А», значить, африканських. Було оголошено, що це не узгоджується з еволюцією жіночих, мтДНК, у яких (нині) неафриканські лінії теж виходять з Африки, а чоловічі і жіночі гаплогрупи мали виходити з Африки разом.
Насправді, ці всі заперечення були в принципі невірними. Опоненти або не хотіли, або не могли розібратися, і, як зазвичай, намагалися «брати горлом». Давайте подивимося і переконаємося, що ніяких протиріч насправді немає, і опоненти просто повторювали завчені мантри, що в популяційної генетики занадто часто прийнято.
Недавня книга «Еволюційна генетика людини» -
правильні дані, невірна інтерпретація
Розкриваємо нову книгу - «Еволюційна генетика людини» (Human Evolutionary Genetics), автори Jobling, Hollox, Hurles, Kivisild, Tyler-Smith, рік видання 2014 (саме так, видавництво забігла на півроку вперед), глава 9 - «Походження сучасної людини», стор 304-305. Розділ «Мітохондріальні ДНК». Цитата: «Дослідження показали вражаючі особливості: повне розділення африканських і неафриканських ДНК-ліній» («Complete separation of African and non - African lineages»). Розділ «Y-хромосоми». Цитата: «Хоча менш детальні, ніж мтДНК, дослідження показали близькі паралелі: повне розділення африканських і неафриканських ДНК-ліній» («Complete separation of African and non - African lineages»).
Як ми бачимо, ніяких протиріч з діаграмою вище. Але автори вже накручують свої інтерпретації, грунтуючись на даних 2000 року - і по мтДНК, і по Y-хромосомі. Так, Y-хромосомну гаплогрупи В вважають африканською, і пишуть, що відповідна гілка містить «як африканські, так і неафриканські ДНК-лінії». Дивимося на діаграму - так, гаплогруппа В знаходиться в одному кущі з неафриканськими гаплогрупами, і ми вище показали, що вона віддалена від африканських гаплогрупп, і знаходиться в одному кластері з неафриканськими, з одним загальним пращуром. А чому автори назвали її «африканською»? Та так, багато носіїв гаплогрупи В зараз живуть в Африці. Пам'ятаєте, як я писав про популяційних генетиків? «Що бачу, те й співаю». У них раз в одному кластері знаходяться як гаплогруппа В, так і неафриканські лінії, значить, «вихід з Африки». А вони всі там, в тому кластері, неафриканські. Та якби й була неафриканська лінія разом з неафриканською - чому неодмінно «вихід з Африки»? Чому з тим же успіхом не «вхід в Африку»? А так, марширують по накатаному тракту, відповідь заздалегідь відома. Датування цього кластеру, що містить неафриканські гаплогрупи і гаплогрупи В (теж неафриканську за походженням) автори книги дають як 52±28 тисяч років тому. У мене в статті - 64±6 тисяч років тому. Де ж протиріччя?
Ті ж автори наводять датування всіх ДНК-ліній - 172±50 тисяч років тому. Дійсно, у мене в статті 160±12 тисяч років тому. Де ж протиріччя? Тобто популяційні генетики оскаржують не по суті, не з даними в руках, а просто «в принципі», заради неприйняття. Звичайна справа.
Відносно мтДНК автори симетрично призводять ті ж самі інтерпретації, що і з Y-хромосомою - схожа гілка, що містить «африканські мтДНК» (тому що зараз там живуть) і неафриканські - значить, «вихід з Африки», причому датування цієї «змішаної» гілки між 31 і 79 тисяч років тому, з медіаною 40 тисяч років тому, датування сукупності всіх мтДНК - між 40 і 140 тисяч років тому, медіана 59 тисяч років тому. Різнобій у датуваннях між Y-хромосомними даними і мтДНК автори і не обговорюють, а навіщо? Висновок давно готовий - «вихід людства з Африки». Цей же висновок, хоча в обережному вигляді - і в ув'язненні по чолі. Там же і про «більш високу генетичну різноманітність в Африці», і те, що в Африці людина з'явилася приблизно 200 тисяч років тому, а поза Африкою - після 45 тисяч років тому. Тут же і про консенсус фахівців про «вихід з Африки». Ми бачили, що всі ці (або подібні) датування і «розмаїття» пояснюються діаграмою вище, але попгенетики інших пояснень не хочуть. У них - «консенсус».
Історія з продовженням статті Канн (1987)
«Про вихід з Африки», але вже без Канн (1991)
По-своєму цікаве продовження статті Канн та інших (Cann, Stoneking and Wilson, 1987), яку ми обговорювали вище. Нова стаття вийшла через чотири роки (Vigilant et al, 1991), в авторах Канн вже немає, але є два колишніх співавтора, Стоункінг і Вілсон, з трьома новими авторами. Стаття 1991 повідомляє, що робота Канн та ін. (1987) зустріла різке неприйняття багатьох фахівців з-за того, що загальний пращур людства нібито жив в Африці, і визнає, що в статті Канн та ін. (1987) було багато слабких ланок. Ці слабкі ланки автори (двоє з яких і були авторами тієї слабкої, за їх визнанням, роботи) перераховують протягом цілого абзацу - там і непрямий метод зіставлення мтДНК, і мала вибірка, причому складається в основному з американців африканського походження, та завідомо непридатний метод «середньої точки», застосований авторами статті 1987 року, і відсутність статистичної обробки отриманих даних, і «неадекватна калібрування» швидкості мутацій в мтДНК, та інші. Інакше кажучи, ця завідомо слабка стаття, за визнанням самих авторів, лягла в основу теорії «виходу з Африки». Але процес вже пішов, тому подальша стаття (1991) мала на меті все-таки виправдати концепцію «виходу з Африки», що неафриканці походять від африканців, і фактично замінити собою слабку, розкритиковану статтю 1987 року.
І в чому було те саме виправдання? Показати, що африканські мтДНК древніші неафриканських. Але це знову продовження тієї вічної фундаментальної помилки популяційних генетиків, що якщо одна популяція древніша іншої - то перша популяція нібито пращурна по відношенню до другої. Погляньмо знову на ту ж саму діаграму вище - ліва гілка давніша правої, але вона не пращурна стосовно правої. У них - один загальний пращур. І ось цю фундаментальну помилку попгенетики повторюють всі наступні 25 років, до теперішнього часу. Знову і знову автори статті (1991) повторюють, що африканська гілка древніша неафриканської, значить, пращурна, не розуміючи, що це зовсім не є доказом «пращурності». Мій дядько «древніше» ніж я, але він не мій пращур.
У висновку статті (1991) автори пишуть: ми представили найбільш суворі докази, що наш спільний пращур жив в Африці 200 тисяч років тому. Насправді, як читач вже давно зрозумів, ці докази насправді були про те, що збережена гілка людей, що нині живуть в Африці, древніша збереженої гілки людей, що живуть поза Африкою. Про «пращурну» ці «докази» нічого не говорять. Для цього треба порівнювати гаплотипи популяцій (що автори статті 1991 року не робили, і попгенетики не роблять досі) та їх сніп-мутації (що автори не робили), і останнє теж показує, що наші пращури з Африки не вийшли. Про це - наступний розділ.
Сніп (SNP) - мутації показують, що ми -
НЕ нащадки африканців гаплогрупп А або В
Перейдемо до недавної статті (Scozzari et al, 2012), яку часто ставлять у приклад як зразкову роботу по геному африканців та обгрунтуванню «виходу людства з Африки». Дійсно, стаття оголошує про виявлення 22 нових необоротних мутацій в Y-хромосомі людини, про підтвердження 146 відомих мутацій і про конструювання нового, поліпшеного дерева гаплогрупп і субкладів африканців з переходом в неафриканську частину дерева, і саме зведеної гаплогрупи СТ. Це - вся права частина дерева вище на діаграмі, від гаплогрупи С до R2. Автори статті називають її «вийшла з Африки». Подивимося, чи це так. Дерево гаплогрупп і субкладів зі статті (Scozzari et al, 2012):
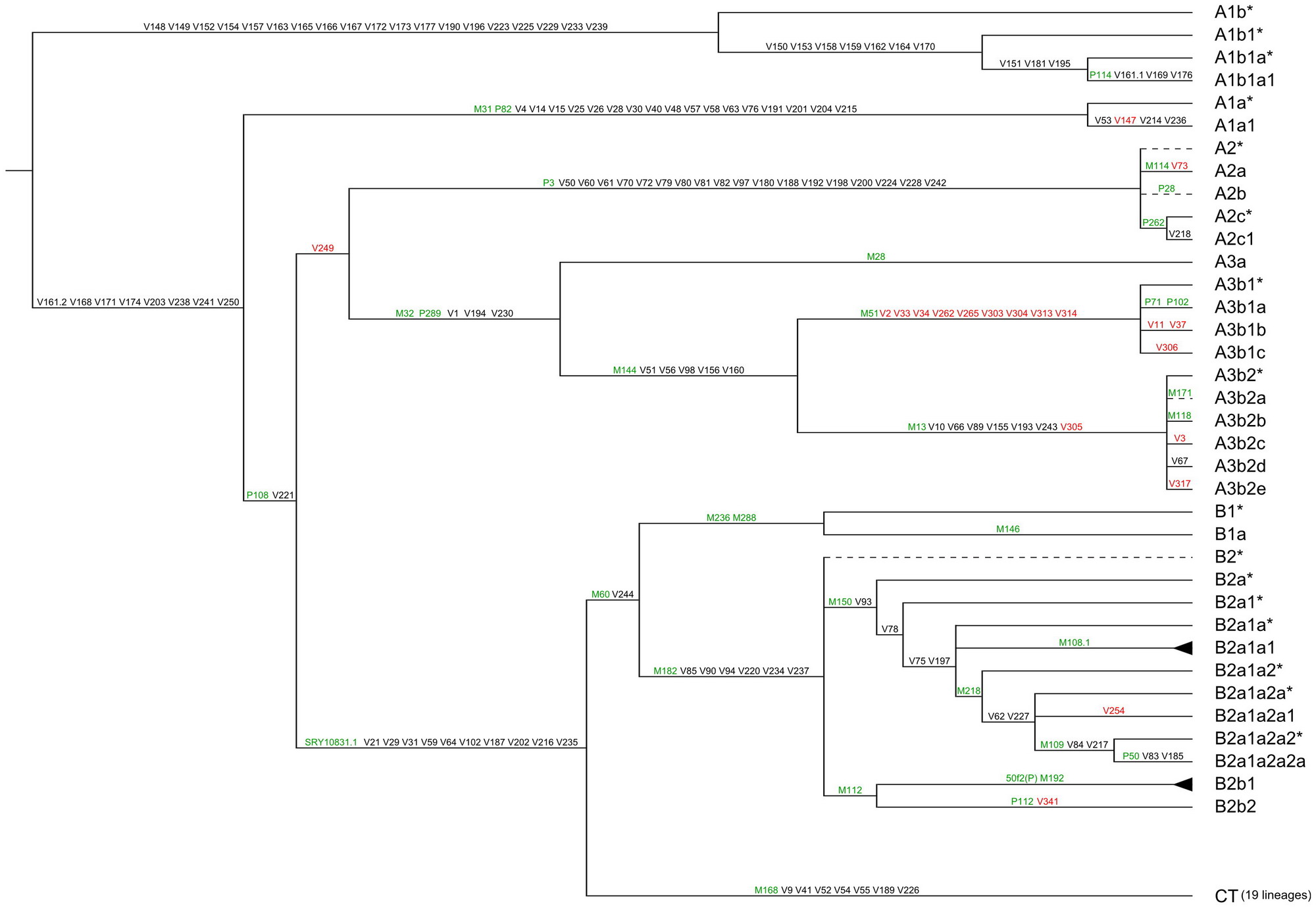
Дерево найдавніших гаплогрупп і субкладів, наведена в статті (Scozzari et al, 2012). Показані номери необоротних мутацій (SNP, Single Nucleotide Polymorphism, або сніпи), які задають певні субклади. Видно, що більше половини субкладів відносяться до гаплогрупи А, яку автори вважають африканською. Всі інші субклади, крім одного, відносяться до гаплогрупи В, яку автори також вважають африканською. Гаплогруппа в нижній правій частині, СТ, складається, за даними авторів, з 19 ДНК-ліній, все неафриканських. Картинка клікабельні.
Звернемо увагу на деякі особливості дерева на цьому малюнку. Воно починається (точніше, продовжує еволюційний стовбур Y-хромосоми людини) у верхній лівій частині діаграми, тут же йде першим розбіжність, або вилка (гаплогруппа A0-T, хоча ця назва на діаграмі не показана), на гаплогрупу A1b (так на діаграмі) c субкладами, з одного боку, і на решту частини дерева, з іншого. Інакше кажучи, від дерева відходить перша африканська гілка, і ніякі неафріканці (гаплогруппа СТ) від неї вже не походить. У статті використовується вже застаріла номенклатура 2011 року, і те, що в статті позначено як A1b, зараз називається А0, з сніпами V148, V149 та іншими, наведеними на верхній лінії діаграми (див. вище також дерево гаплогрупи А).
На наступній вилці (гаплогруппа А1) у бік відходять африканські гаплогрупи А1А і А1А1 з їх сніпами М31, Р82, V4 та іншими, в інший бік - інша частина дерева. Від другої африканської гілки (A1a c cубкладом) неафріканці (гаплогруппа СТ) теж не відбуваються.
Третя вілка - гаплогруппа А1b згідно з нинішньою класифікацією. Від неї відходять убік африканські гаплогрупи А2 і А3 з субкладами (номенклатура застаріла), зараз це A1b1 з сніпом V249/L419, яка далі розходиться на субклади A1b1a-V50 (колишня А2) і A1b1b-M32 (колишня А3), обидва з підгрупами. Серед останніх - субклад A1b1b2b-M13, той самий, з якого виходять арабська та європейська ДНК-лінії, гаплотипи яких ми розглядали вище. Інша гілка цієї вілки від гаплогрупи А1b - зведена гаплогруппа ВТ, показана в нижній частині малюнка зі статті (Scozzari et al, 2012). Абсолютно ясно, що ця гаплогруппа ВТ жодним чином не утворюється від «африканських» гаплогрупп з індексом А, які всі знаходяться у верхній частині цього малюнка. Слово «африканські» тут доводиться брати в лапки, оскільки серед їх субкладів і ті самі європейські і арабські гілки, причому європейська - це, в основному, Англія, Ірландія, Шотландія, Туреччина (хоча лише 3% Туреччини територіально знаходиться у Європі), арабська - в основному Саудівська Аравія, і гаплотипи з Англії, Швейцарії, Фінляндії та інших країн.
Природно, можна стверджувати, що європейські та азіатські гаплотипи гаплогрупи А колись вийшли з Африки разом з мігрантами звідти, але з таким же успіхом можна стверджувати, що вони потрапили в Африку тим же шляхом. Так що ці аргументи не проходять, хоча тільки «з Африки» викликає підтримку серед прихильників «виходу з Африки». Альтернативні пояснення вони не розглядають в принципі.
Завершуючи розгляд малюнка зі статті (Scozzari et al, 2012), варто відзначити і той факт, що загальновизнано «неафриканських» зведена гаплогруппа СТ (нижня лінія на малюнку) не виходить і з гаплогрупи В з її субкладами, назвати її африканською чи ні. Видно, що еволюційний шлях гаплогрупи СТ мине всі «африканські» гаплогрупи, навіть якщо вони всі містять європейських чи інших пращурів. Якщо рухатися вгору по часовій шкалі (тобто зліва направо) мутаційний шлях гаплогрупи СТ виходить з основного "стовбура" еволюції Y-хромосоми людини, тобто нижче «хвостика», позначеного на малюнку зліва вгорі, проходить через гаплогрупи А0-Т (ніяких відомостей, що вона нібито «африканська» - ні), далі через гаплогрупи А1 (те ж саме, немає ніяких відомостей, що вона нібито «африканська»), далі через гаплогрупи A1b, теж неафриканських, далі через гаплогрупи ВТ, і стає гаплогрупою СТ. Всі три «африканські» гілки (А0, А1А, А1b1) на цьому шляху йдуть у бік шляхом відповідних відгалужень, гілок.
Ми зупинилися на цьому настільки детально, тому що в академічній літературі, і тим більше в популярній, такий чи подібний аналіз ніколи не проводився. Зазвичай показується дерево, як на малюнку зі статті (Scozzari et al, 2012), і скоромовкою повідомляється, що воно «свідчить про вихід людства з Африки». Ніякого пояснення не дається. Відповідь готова вже заздалегідь. Іноді доходить до абсурду - повідомляється, що гаплогрупи ВТ і СТ виходять з гаплогрупп А1b, або A1, або А0-Т, значить, це африканські гаплогрупи, оскільки мають індекс «А». Тобто підміна понять зайшла настільки далеко, що абсолютно умовні назви приймаються за доведення тези. Те, що ці гаплогрупи могли з таким же успіхом назвати X, Y, Z, або W, в голову вже не приходить. Раз «А», значить, Африка, немає і сумнівів.
Так що ніякого протиріччя між малюнком зі статті (Scozzari et al, 2012) і діаграмою вище немає, вони показують ті ж самі закономірності еволюційного розвитку гаплогрупп, а саме розбіжність на «африканські» і «неафриканські» гілки. Різниця між ними тільки в тому що на малюнку зі статті (Scozzari et al, 2012) більш детально показані субклади гаплогрупп А («африканської») і В, а на діаграмі - «неафриканські» гаплогрупи ВТ. Ще в тому, що перше зображення побудовано з урахуванням хронологічної шкали, а друга - без такого врахування. Щоб показати схожість, розташуємо обидва дерева гаплогрупп вертикально, в одному напрямку.
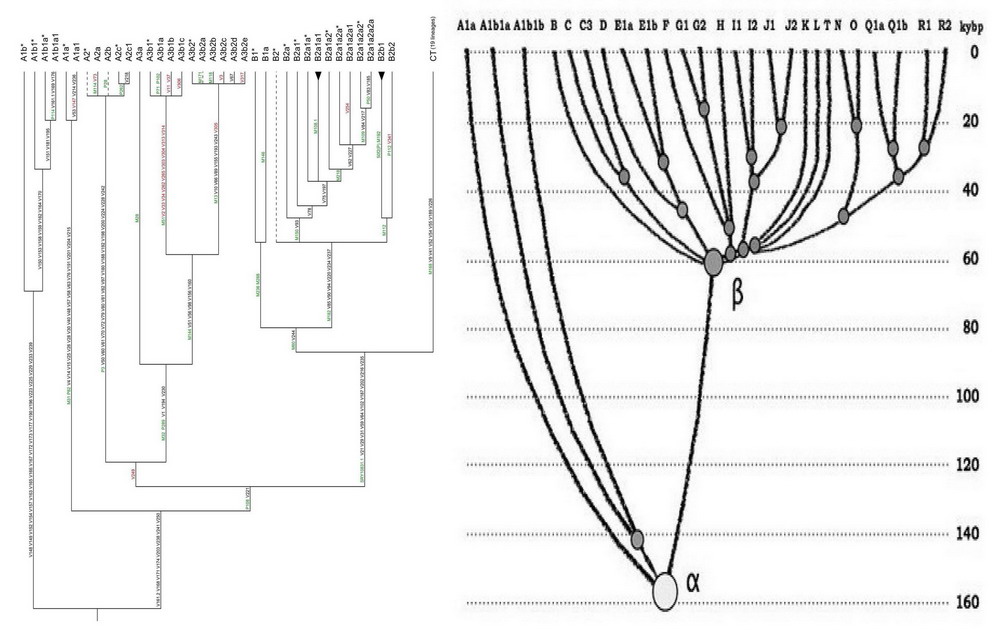
Зіставлення дерев гаплогрупп і субкладів «африканської» групи (ліва частина обох дерев) і «неафриканських» (одна лінія зведеної гаплогрупи СТ на лівому дереві і кущ гаплогрупп ВТ на правому дереві). Ліве дерево (Scozzari et al) опубліковано в листопаді 2012 року, праве дерево (Klyosov, Rozanskii) опубліковано в травні 2012 року. Ліве дерево показує, що серія «африканських» гаплогрупп тричі послідовно відокремлюється і йде в бік від неафриканських, і що неафриканська ДНК-лінія СТ (вертикальна лінія праворуч) не відбувається від «африканських». Праве дерево показує ті ж відщеплення «африканських» гаплогрупп від неафриканських (кущ гаплогрупп праворуч), і вказує, що розбіжність дерева на африканських та неафриканських гаплогруппа походить приблизно 160 тисяч років тому. Картинка клікабельні.
Обидва дерева не показують недавно виявлену гаплогрупи А00, «вік» якої не менше 200 тисяч років. Вона наведена на наступному малюнку, разом із змінами номенклатури (згідно ISOGG, 2013).
Отже, картина в цілому прояснилася. Ніякого протиріччя між деревом Y-хромосомних гаплогрупп людства, отриманим при вивченні гаплотипів гаплогрупп від А до Т (Klyosov and Rozhanskii, 2012, травень 2012), і деревом, отриманим при геномному вивченні Y-хромосоми (Scozzari et al, 2012, листопад 2012), немає. Всі ці дані, як і інші, показують глибокі мутаційні розбіжності між африканськими і неафриканськими гілками (гаплогрупами, субкладами), і не виявляють «африканське» походження анатомічно сучасного людства. Замість цього, дані показують розбіжність африканських і неафриканських ДНК-ліній приблизно 160 тисяч років тому.
Виникає закономірне і здивоване запитання - чому, маючи всі ці дані, автори досліджень продовжують писати, що людство вийшло з Африки відносно недавно, в останні 50-100 тисяч років? На якому рівні фактичного матеріалу або його інтерпретацій відбувається збій? Відповісти на це питання видається не менш важливим, ніж отримати відповідь, що людство з Африки не виходило.
Подивимося на згадану статтю Scozzari et al (2012). У який момент там з'явилася фраза про африканське походження людини? На чому заснована?
Ця фраза з'являється вже в другому абзаці введення в статтю, і повідомляє, що гаплогруппа СТ є результатом «недавнього виходу з Африки». У підтвержденні цього дається посилання на статтю консорціуму Проекту «1000 геномів» під назвою «Карта варіацій генома людини» (Nature, 2010), в якій про вихід з Африки взагалі ні слова, як і про гаплогрупи СТ. Розумієте, в чому проблема? Прихильників концепції «виходу з Африки» доводиться постійно ловити за руку, і це триває вже більше 20 років. Ще кілька абзаців нижче - знову про «вихід з Африки», вже гаплогрупи С, і взагалі ніякого посилання.
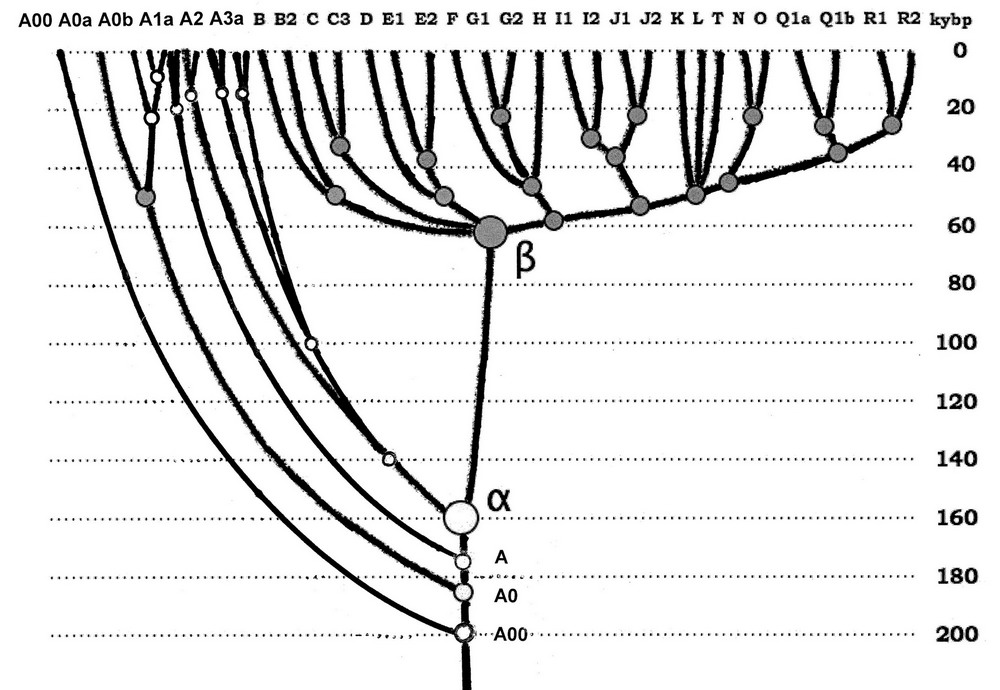
Діаграма еволюції гаплогрупп сучасного людства з додаванням недавно виявленої гаплогрупи А00, і з заміною застарілої номенклатури 2012 року на номенклатуру 2013 року. На горизонтальній осі - основні гаплогрупи Y-хромосоми людства, на вертикальній - абсолютна шкала часу. Спільний пращур альфа-гаплогрупи (гаплогрупи A1b в нинішній класифікації) жив приблизно 160 тисяч років тому, загальний пращур бета-гаплогрупи (або пращурний для гаплогрупп від В до Т) - 64±6 тисяч років тому (зі статті AA Klyosov and IL Rozhanskiі, Advances in Anthropology, 2012b).
Йдемо за статтею (Scozzari et al, 2012) далі. Описується гаплогруппа A1b (сама верхня лінія на малюнку, взятому з цієї статті, яка першою відійшла від решти частини дерева, і за новою номенклатурою називається гаплогрупою A0). Вона ж відходить вліво від дерева на оновленій діаграмі еволюції гаплогрупп сучасного людства приблизно 180 тисяч років тому. Повідомляється, що людей з такою мутацією (Р114) знайдено дуже мало, всього троє з Камеруну, з них один - в даній роботі. Мій коментар - дуже добре, у мене особисто немає великих сумнівів, що гаплогруппа А0 і її гілки - африканські. Але ми від них не походимо, що дерево і показує.
Далі, автори повідомляють, що в Нігері знайшли двох людей гаплогрупи А1А - друга лінія у верхній частині малюнка зі статті (Scozzari et al, 2012), теж «африканська». Мій коментар - і з цим немає жодних проблем. Неафриканських нащадків від них теж немає, згідно тому ж малюнку.
Далі, гаплогруппа А2, тобто за нинішньою номенклатурою А1b1a, третя за рахунком лінія на малюнку зі статті. Автори повідомляють, що носії цієї гаплогрупи майже всі говорять на клацаючих мовах південної Африки, а також є пігмеями центральної Африки. Автори знайшли трьох носіїв цієї гаплогрупи в Південній Африці. Мій коментар - чудово, немає проблем, це цілком африканська лінія, і неафріканці від неї не походять, як показує той же малюнок.
Відносно гаплогрупи А3, тобто за нинішньою номенклатурі A1b1b-M13, автори знайшли десять носіїв цієї гаплогрупи - в Ефіопії, Кенії і Південній Африці. Ще 28 осіб, володарів цієї гаплогрупи з європейських країн і Саудівської Аравії, наведені у Проекті гаплогрупи А, на який вище дан лінк. Але нехай ця гаплогруппа вважається африканською, все одно неафріканці від неї не походять, як видно з малюнка статті (Scozzari et al, 2012). Гаплогрупи В автори відправляють на південь від Сахари, а також по всій Африці - центральної, східної і південної. Ми вже показували вище, що гаплотипи гаплогрупи В надзвичайно віддалені від «африканських», і явно мають інше походження, споріднене неафриканським гаплогрупам. Але це в даному випадку не має значення, так як «неафриканські» ДНК-лінії гаплогрупи СТ не виходять з гаплогрупи В. Вони з нею мають спільного пращура - гаплогрупи ВТ.
І який же після всього цього роблять висновок автори статті (Scozzari et al, 2012)? Ви, читачу, будете сміятися, але висновок такий, що гаплогрупи СТ вийшли з Африки, і не тільки з Африки взагалі, а з її північно-західного регіону. Саме там, за словами авторів, знаходяться «витоки різноманітності Y-хромосоми людства». Як, звідки? Адже навіть побіжний погляд на дерево гаплотипів показує, що в основі СТ немає африканських джерел. Всі вони - у верхній частині діаграми. А ось так. На цей рахунок у Америці є приказка - «не плутайте мене фактами, я вже налаштувався».
Як читач уже зрозумів, і зрозумів давно, «вихід людства з Африки» став форменною релігією, заснованої, як і належить релігії, на вірі, і аргументи там вже майже марні.
Про це ж і інша відносно недавня стаття (Cruciani et al, 2011), в якій вже в назві варто «походження різноманітності в Африці». На якій науковій підставі? Та все на тому ж - показали, що африканські Y-хромосомні лінії древніші, ніж лінії неафриканців. Знову відповідно до діаграми вище. Їх дерево гаплогрупп майже таке ж, як і на малюнку зі статті (Scozzari et al, 2012), але з датуваннями - 142 тисяч років тому в сторону від еволюційного Y-хромосомного стовбура відійшла гілка А1b (A0 в новій класифікації), потім приблизно 108 тисяч років тому відійшла гілка А1А, потім, 105 тисяч років тому, гілка А2, потім, ті ж 105 тисяч років тому, гілка А3, які автори вважають африканськими, оскільки вони знайдені у чотирьох африканців, у яких ці гаплогрупи визначали - і нехай вважають, і тільки потім, 75 тисяч років тому, відійшла гілка ВТ і далі, 39 тисяч років тому, гілка СТ, вже загальновизнано неафриканською. Ні ВТ, ні СТ від «африканських» ліній не походять. Але оскільки точки розгалуження авторами названі буквою «А» (А1А-Т, А2-Т), що автоматично приймається за «африканські», то ось і «походження з Африки». Вражає такий тип прямолінійного мислення популяційних генетиків.
Добре, це - чоловіча, Y-хромосома. Ясно, що відповідні експериментальні дані не показують ніякого виходу з Африки. Немає в основі неафриканських гаплогрупп африканських сніп-мутацій. Неафриканські гаплотипи виключно віддалені від африканських. Як визнають практично всі джерела, між ними - величезний розрив, але автори далі не йдуть. Закривши очі, повторюють як мантру - «вийшли з Африки».
Перехресна перевірка діаграми за допомогою сніпів (SNP)
Існує ще один підхід до перевірки топології дерева, представленого на діаграмі трохи вище, за допомогою сніпів. Справа в тому, що в ході еволюційного розвитку Y-хромосоми в ній накопичуються практично незворотні мутації, так звані SNP (Single Nucleotide Polymorphism), або сніпи. Чим довший перехід в діаграмі, тим більша вірогідність сніпа, тим їх більше накопичується в Y-хромосомі (і в інших хромосомах, але в даному випадку ми розглядаємо тільки Y-хромосому. Найпротяжніша еволюційна лінія - гаплогрупи А00, вона сама архаїчна, тому в носіях цієї гаплогрупи повинна виявитися максимальна кількість сніпів. На другому місці за протяжністю - гаплогруппа А0, на третьому - лінії гаплогрупи А (А1А). Відстань між альфа- і бета-гаплогрупи (тобто від А1b до ВТ) повинно бути відносно невеликим (у порівнянні з протяжністю ліній А00 і А0), і далі сніпи вже накопичуються при переході від ВТ по лініях конкретних, вже більш сучасних гаплогрупп.
Так і виявилося, на підтвердження діаграми. Я зараз наведу список сніпів по кожній з перерахованих гаплогрупп - з одного боку, створивши якийсь компактний довідник, щоб з ним можна було працювати, кому потрібно, а з іншого, щоб не просто обмежитися на словах якимось числом, яке перевірити важко. Правда, ці числа сніпів не є остаточними - час від часу відкривають нові сніпи. Далі, оскільки сніпи з'являються неупорядкованно, то ми маємо справу зі статистикою, а не з абсолютними, непорушними числами. Так що список нижче і число сніпів по кожній гаплогруппі знаходяться в розвитку, хоча загальне уявлення дають.
Отже - гаплогруппа А00, сама древня, сама архаїчна, її еволюційна лінія на діаграмі найдовша. Ці сніпи виявили в племені Mbo (Мбо, російськими буквами) в африканському Камеруні:
AF4, AF5, AF7, AF8, AF9, AF10, AF13, L990, L1086, L1087, L1088, L1091, L1092, L1094, L1096, L1097, L1100, L1102, L1103, L1104, L1106, L1107, L1108, L1109, L1110, L1111, L1113, L1114, L1115, L1117, L1119, L1122, L1126, L1131, L1133, L1134, L1138, L1139, L1140, L1141, L1144, L1146, L1147, L1148, L1149, L1151, L1152, L1154, L1156, L1157, L1158, L1159, L1160, L1161, L1163, L1233, L1234, L1236, L1284.
Всього в гаплогруппі А00 - 59 сніпів. Оскільки вік гаплогрупи А00 оцінюється приблизно в 210 тисяч років, то звідси можна припустити, що сніп - мутація відбувається в середньому раз в 3600 років.
Далі, як ми вже знаємо, на стовбурі дерева гаплогрупп (діаграма вище) з'явилася гаплогруппа А0-Т, від неї відійшли гаплогрупи А0 і А1, остання продовжила стовбур дерева. А0 зараз живуть в основному в Африці. Жодної сніп - мутації зі списку А00 у А0-Т немає. Тобто А0-Т не походить від африканської гаплогрупи А00. Немає підстав вважати гаплогрупи А0-Т африканською. Але вона - пращур африканської лінії А0, і нас, неафриканців (пращури яких пройшли через гаплогрупи ВТ, далі СТ, і так далі).
Сніпів гаплогрупи А0-Т, їх 32:
AF3, L1085, L1089, L1090, L1093, L1095, L1098, L1099, L1101, L1105, L1116, L1118, L1120, L1121, L1123, L1124, L1125, L1127, L1128, L1129, L1130, L1132, L1135, L1136, L1137, L1142, L1143, L1145, L1150, L1155, L1235, L1273
Гаплогруппа А0 має наступні сніпи, їх 51:
L529.2, L896, L982, L984, L990, L991, L993, L995, L996, L997, L998, L999, L1000, L1001, L1006, L1008, L1010, L1011, L1012, L1015, L1016, L1017, L1018, L1055, L1073, L1075, L1076, L1077, L1078, L1080, V148, V149, V152, V154, V157, V163, V164, V165, V166, V167, V172, V173, V176, V177, V190, V196, V223, V225, V229, V233, V239
Як видно, в гаплогруппі А0 на 8 сніпів менше, ніж в А00, тобто вона приблизно на 30 тисяч років молодша. І дійсно, вік гаплогрупи А0 оцінюють в 180 тисяч років, на 30 тисяч років менше, ніж вік гаплогрупи А00.
У гаплогруппі А1 поки виявили 21 сніп:
L985, L986, L989, L1002, L1003, L1004, L1005, L1009, L1013, L1053, L1084, L1112, L1153, P305, V161.2, V168, V171, V174, V238, V241, V250
На відміну від братньої гаплогрупи А0, носії А1, мабуть, не дожили до теперішнього часу. Можливо, вони і потрапили в той катаклізм, який привів до пляшкового горлечка неафриканських популяцій світу. Для оцінки сумарного часу життя гаплогрупп А0-Т, А1 і A1b (в останної - всього дві сніп-мутації, Р108 і V221) слід скласти число їх сніп-мутацій, виходить 32+21+2 = 55 мутацій, що приблизно еквівалентно 198 тисяч років.
Нарешті, в гаплогруппі ВТ - 30 сніп-мутацій
413, L418, L438, L440, L604, L957, L962, L969, L970, L971, L977, L1060, L1061, L1062, M42, M91, M94, M139, M299, P97, SRY10831.1, V21, V29, V59, V64, V102, V187, V202, V216, V235
Це дає 108 тисяч років еволюції бета-гаплогрупи від часу розбіжності з африканською гаплогрупою A1b1 (160±12 тисяч років тому) до проходження пляшкового горлечка популяції 64±6 тисяч років тому. Це і є зниклі 108 тисяч років на діаграмі (дистанція між альфа- і бета-гаплогрупою).
Важливо те, що ці зниклі тисячоліття в еволюції неафриканських ДНК-генеалогічних ліній відтворюються як при аналізі гаплотипів (на підставі чого і була побудована діаграма вище), так і при аналізі сніп-мутацій. Ось це і є перехресна перевірка діаграми. Чому ми не бачимо цих зниклих людей в Євразії - невідомо. З іншого боку, відомі скелетні залишки Homo sapiens з датуванням між 160 і 60 тисяч років, виявлені на Близькому Сході, але їх гаплогруппа не перевірялася. Якщо виявиться гаплогруппа BT з якоюсь з мутацій, наведених у списку вище - загадка буде остаточно вирішена.:
Неафриканські мтДНК не походять з африканських
А що на цей рахунок говорять дані по жіночій, мтДНК? Ну, кажуть прихильники «виходу з Африки», там-то тим більше ясно. Неафриканські мтДНК «всі походять з африканських». Чи так це?
Погляньмо на недавню статтю відомого генетика Д. Бехара (Behar et al, 2012), в якій була проведена принципова ревізія характеру уявлення послідовності мтДНК. На наступному малюнку видно, що на самому початку еволюційного дерева мтДНК «сучасної людини» відбувається розходження гаплогрупп на L0 (гілка праворуч) і гаплогрупи L1-L6 (гілка ліворуч), з яких далі відбуваються всі наступні гаплогрупи. Гаплогруппа L0 насправді представляє велику серію з півсотні стародавніх африканських гаплогрупп, в основному знайдених на південь від Сахари - серед койсанського населення Південної Африки, а також в Ефіопії і Танзанії (Східна Африка), у Мозамбіку (Південно-Східна Африка), і серед пігмеїв. Всі інші мтДНК походять, як прийнято більшістю фахівців, з гаплогрупи L3, вік якої за оцінками приблизно 60-70 тисяч років, тобто такий же, як гаплогрупи ВТ Y-хромосоми. Тобто цілком імовірно, що гаплогруппа L3 не вийшла з Африки, а, навпаки, прийшла в Африку разом з носіями Y-хромосоми, наприклад, гаплогрупи ВТ. Який же висновок робить Бехар з співавторами? Природно, «людство вийшло з Африки». Глибоке розходження африканської гаплогрупи L0 з іншими в їхній статті навіть не обговорюється, хоча з даних статті відразу видно, що африканська гаплогруппа L0 не є пращурною по відношенню до всіх інших гаплогруп мтДНК.
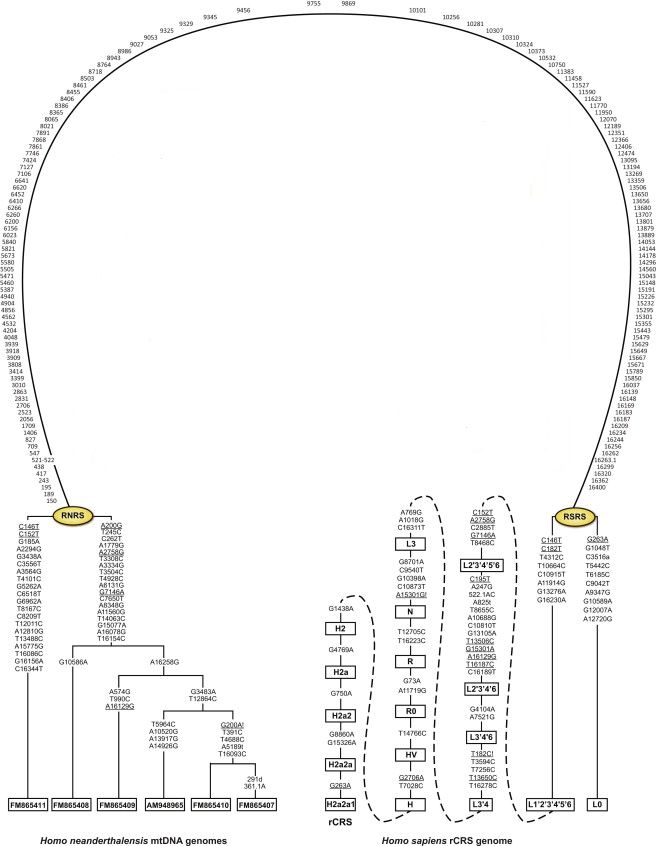
Схематичне представлення мтДНК людини, що показує мутаційнний зв'язок між мтДНК неандертальця (ліворуч) і мтДНК людини розумної (справа). У овалі ліворуч і праворуч скорочення RNRS і RSRS означають відповідно «реконструйована референсна послідовність неандертальця» і «реконструйована референсна послідовність людини». З роботи (Behar et al., 2012). Зверніть увагу на різке розходження на початку референсної послідовності людини (праворуч) ланцюга мутацій на африканську гаплогрупи L0 (справа внизу), і ланцюжок всіх інших гаплогрупп мтДНК. Картинка клікабельні.
Насправді, розбіжність африканської гаплогрупи L0 (вік 150-170 тисяч років) і спочатку неафриканських гаплогрупп L1-L6, з подальшим приходом гаплогрупи L3 (вік 60-70 тисяч років) до Африки виявляється практично збігається з розбіжністю Y-хромосомних гаплогрупп африканської серії А (вік 160 тисяч років) і спочатку неафриканських ВТ (вік 64 тисячі років ) з наступним приходом гаплогрупи В в Африку.
Так що і відносно мтДНК концепція «виходу з Африки» виявляється абсолютно необов'язковою, і фактично побудована на піску. Опис мтДНК в академічних статтях і довідниках рясніє словами «можливо», «мабуть», «передбачається», що фактично означає, що даних немає, і що все побудовано на припущеннях. Одна проблема - всі ці припущення завжди трактуються тільки в одну сторону - «виходу з Африки».
Разом з тим накопичується все більше даних, що в древньому минулому міграції популяцій в Африку відбувалися багато разів. Щойно вийшла стаття в журналі Nature (Hayden, 2013), в якій повідомляється про дві міграції з Євразії в койсанські племена в Південній Африці, одна 3 тисячі років тому в східну Африку, інша, її продовження - 900-1800 років тому в Південну Африку. Які гаплогрупи принесли мігранти - не повідомляється. Але немає сумніву, що вони різко збільшили «генетичну різноманітність» койсан, яке вважається максимальним в Африці. Друга важлива особливість цього повідомлення - воно вказує на можливість міграцій «в Африку», у чому, втім, ніякого сумніву і не могло бути. Неясно, чому з таким завзяттям прихильники « виходу людства з Африки» тримаються саме за їх односторонній варіант. Втім, завзятість зменшується, і ось вже Сара Тишкоф з Пенсільванського університету, одна з найбільш активних захисників «з Африки», вже вітає нові дані «в Африку», і називає їх «мають сенс», оскільки «про це говорять і археологічні, і лінгвістичні дослідження» (Nature, 29 серпня 2013, стор 514).
Міграції вірусу герпесу помилково оголошені як
нібито підтверджують «вихід людства з Африки»
Історія з нещодавньої нашумівшої публікації «Використання філогенетики генома простого вірусу герпесу для простеження древніх міграцій людини» є показовою - як вихідна установка на нібито вихід сучасного людства з Африки спотворює висновки наукової публікації. Нагадаю, що в статті зіставляли 31 геномні послідовності простого вірусу герпесу в популяціях зі Східної Африки (Кенія), Східної Азії (Китай, Південна Корея, Японія), Північної Америки (США) і Європи (Сполучене Королівство), і виявили, що побудоване філогенетичне дерево розходиться на шість кластерів. Ці кластери відповідали наступним популяціям:
I - з 10 зразків 7 були з Сіетла, один «із США», один із Сан-Франциско, і самий нижній - (номер 17) з Шотландії.
II - з 6 зразків один був з Китаю, один з США (Х'юстон), два з Південної Кореї і два з Японії.
III - два зразка з Кенії.
IV - три зразка з Кенії.
V - сім зразків з Кенії.
VI - два зразка з Кенії.
Автори статті дані «округлили» і повідомили, що перший кластер «об'єднує Північну Америку та Європу», другий - Східну Азію, решта - «Східну Африку». Походження американських зразків вірусу в статті не дано, тобто незрозуміло, отримані вони від американських індіанців або, наприклад, нащадків англосаксів, але оскільки автори припустили, що випадає з навіть округленого варіанту зразок з Х'юстона, який потрапив до східно-азіатським вірусам, міг належати американському індіанцеві, то стає ясно, що інші північно-американські зразки - швидше за все від нащадків європейців. Проте, автори зробили висновок, що ці кластери відображають «глобальні міграції (стародавньої) людини», і підтверджують вихід сучасного людства з Африки (supports... the «out of Africa» theory of human evolution). Основна причина - як звичайно, аргумент «розмаїття», а саме що «віруси східно-африканського походження мають найбільшу різноманітність і утворюють чотири з шести кластерів».
Давайте, як ми вже робили вище, подивимося, звідки з'являється ця «різноманітність». А з'являється воно знову від розбіжності популяцій у різні сторони - одні йдуть в Африку, інші не йдуть. Але вони знову не відбуваються один від одного, вони щоразу походять від більш давнього загального пркщура, який в Африці цілком міг не жити. Тобто знову повторюються ситуації, описані раніше в цій статті. Ніякого «африканського походження» показані дані не дають.
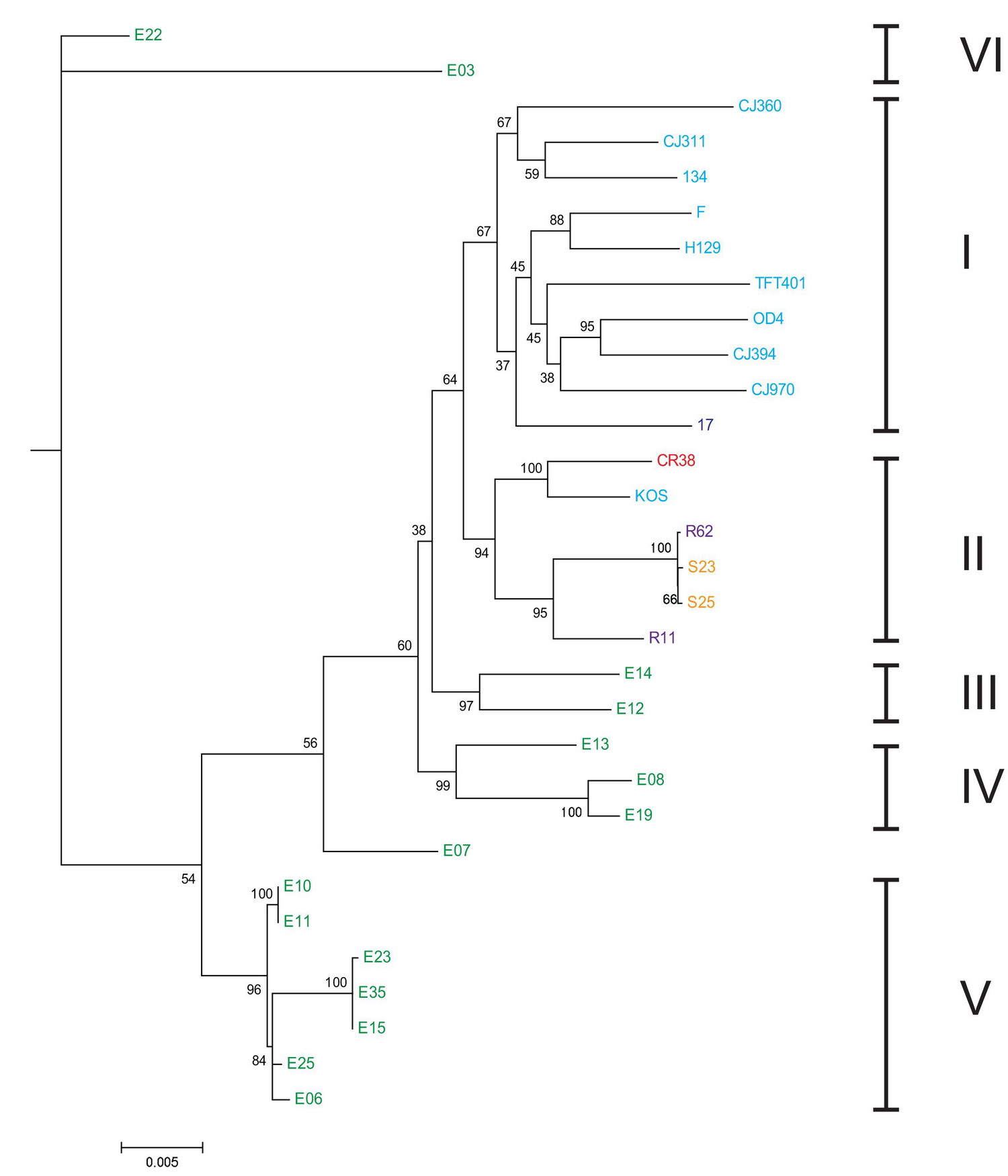
Філогенетичне дерево, що показує динаміку штамів простого вірусу герпесу HSV-1 і його каскадне розходження на шість гілок, як нараховують у роботі (Kolb et al., 2013), які, на думку тих же авторів, слідують певній географії: гілка I - Європа/Північна Америка (штам 17 - з Глазго, решта - Сіетл, Сан Франциско, «США»), гілка 2 - Східна Азія (Китай, Південна Корея, Японія, і Х'юстон [CША]), гілки III, IV, V і VI - східно-африканські (всі з Кенії).
Подивимося на наведену діаграму. Зліва - перше розходження від стародавнього загального пращура. У бік відходять віруси, які сьогодні знаходять у кенійців (два самих верхніх зразка, кластер VI). Від них ніякі неафриканські віруси не походять.
Далі - наступні розходження популяцій вірусів (і, мабуть, їх носіїв - людей). У бік відходить кластер V, нині кенійці. Від них ніякі неафриканські віруси не походять.
Далі - знову вілка, чергове розходження популяцій. У бік відходить знову кенійський вірус, з індексом Е07 на схемі вище. Від нього неафриканські віруси знову не походять. Його об'єднання з іншими кенійськими зразками в кластер IV - помилка авторів, немає там загального кластера.
Слідом - ще дві вілки-розходження популяцій, і кожен раз в сторону відходить кенійська група (кластери IV і III), від якої європейці і азіати не походять. При останньому розходженні в сторону відходить виключно неафриканська група. Вона знову не походить від африканців.
Останнє розходження - на європейські і східно-азіатські (за деяким винятком) лінії вірусів герпесу, кластери I і II. Америка там, судячи з усього, не при чому, це не древні мешканці Америки, а відносно недавні переселенці з Європи.
Вражаюче, що автори не бачать такого простого пояснення динаміки вірусів. Вони повністю поглинені тим, як підігнати дані до «виходу з Африки». Навіщо, яка мета - зрозуміти неможливо.
У своєму коментарі до статті, провідний автор Куртіс Брандт, професор медичної мікробіології та офтальмології, повідомив, що результати виявилися «приголомшуючими», і далі - «ми знайшли, що всі африканські ізоляти утворюють один кластер, всі віруси з Далекого Сходу, Кореї, Японії, Китаю разом утворюють інший кластер, і всі віруси з Європи та Америки, за одним винятком, утворюють ще один кластер». Загалом, незважаючи на очевидні натяжки, це вірно. Але чи показує це «африканське походження сучасної людини»? Ясно, що ні, як це пояснено вище.
В іншому коментарі д-р Брандт поділився цілком відверто: «ми знайшли точно те, що нам сказали молекулярні генетики, які вивчають геном людини, а саме звідки люди походять (з Африки - АК) і як вони розходилися по планеті». Ось це і є основна проблема цього і подібних досліджень. Їм сказали (невірно), вони взяли як керівництво до дії і знайшли саме це.
Стаття надає ще одну ілюстрацію того, як дані підганяються під бажаний «вихід з Африки». Це те, як автори розраховували часи цього «виходу», базуючись на тих самих передбачуваних 50 тис. років тому, і насправді ніколи не показаних. Це - «консенсусні дані». Насправді, різні автори наводять датування від 27 до 200 тисяч років тому, як правило, знову ж таки без розрахунків, але цифри 50 або 70 тис. років тому чомусь виглядають їм привабливими. Нещодавно, правда, «консенсус» почав зміщуватися в 100-140 тис. років тому, але 50 або 70 тис. років за інерцією продовжують наводитися. Ось і автори обговорюваної статті їх, ці умоглядні датування, використовували як базові. Давайте подивимося, що з цього автори отримали. Це дуже повчально.
Як повідомили автори статті, в літературі є три істотно розрізняючі величини констант швидкостей мутацій вірусу простого герпесу та інших вірусів герпесу, рівні 3х10-9, 18.2х10-9, і 30х10-9 мутацій на нуклеотид на рік. Застосування їх для розрахунків часів розбіжності вірусу по популяціям людини, описаним вище, дало б розбіжності за часом в 30 разів. Зрозуміло, що така невизначеність авторів не влаштовувала, і вони вирішили фактично підігнати час вихідної розбіжності популяцій під «консенсусний» час «виходу з Африки», і на першому етапі розрахунків підігнати під передбачуваний час розбіжності європейської та азіатської популяції, яке взяли як 23-45 тисяч років тому, процитувавши чотири літературних джерела на цей рахунок. Усереднивши ці величини, автори взяли як «референсну» величину 34,00 ±10,500 років тому. Правда, замість «європейської» автори постійно згадували «європейська/північно-американська», хоча ясно, що по суті це європейська - один зразок вірусу з Шотландії, всі інші (в основному з Сіетла) - швидше за все, нащадки переселенців з Європи. У будь-якому випадку ці цифри занижені, так як поділ гаплотипів Європи та Азії відбулися не пізніше 55-60 тисяч років тому, найдавніші кісткові залишки «анатомічно сучасної людини» в Європі датуються 45 тис. років назазд, аборигени прийшли до Австралії не пізніше 50 тис. років тому, але в статті все настільки приблизне - і датування, і швидкості мутації вірусу, що до цього ніяк не можна ставитися серйозно. Оскільки часи у авторів набагато занижені, то швидкості мутації повинні були вийти набагато завищені. Так і вийшло - підігнана таким чином константа швидкості мутації вірусу виявилася набагато швидше відомих (точніше, літературних) величин, а саме 134х10-9 мутацій на нуклеотид на рік, з верхнім і нижнім межами 214х10-9 і 74.8х10- 9, відповідно.
Використовуючи цю підігнану величину константи швидкості мутації, автори отримали, що вихідне розходження вірусу відбулося 50.3±16.7 тисяч років тому, і постановили, що це відповідає «виходу людства з Африки». Злегка підправлене датування розбіжності європейців і азіатів за розрахунками авторів виявилася 32.8±10.9 тис. років тому, а час розбіжності єдиного китайського зразка і єдиного зразка з Техасу виявилося 15.76±5.3 тисяч років тому, що автори віднесли до заселення Америки, «яке відбулося в той період». Тут коментарі просто зайві.
Все це лягло в основу проголошення авторами, що «вперше показано, що філогенетичні дані по вірусу герпесу підтверджують вихід людства з Африки». Як насправді показано вище, це не має ніякого відношення до «виходу людства з Африки». Більш того, оцінка величини константи швидкості мутації вірусу герпесу, проведена авторами на підставі більш ніж грубих прикидок перевищує три інші літературні величини в 4.5 - 45 разів. Ніяких перехресних перевірочних тестів з верифікації отриманої константи швидкості мутації авторами не робилося. Те, що автори отримали, виходячи з їх константи, що віруси герпесу HSV-1 і HSV-2 розійшлися 2.184±0.753 мільйонів років тому (зверніть увагу на наведену «точність» в три знаки після коми!), Ні про що не говорить, там могло вийти і 20 мільйонів років тому з тим же успіхом, і теж нічого б не сказало - це могло бути у макаки, наприклад. Якщо все-таки більш вірні літературні дані, то вихідне розходження вірусу герпесу могло відбутися не 50.3 тисяч років тому, а 220 тис. років - 2.2 мільйона років тому, і теж могло бути непогано інтерпретовано в рамках еволюції людини. Так що вихідні, експериментальні дані авторів представляють безперечну цінність, а маніпуляції, висновки та інтерпретації - ніякої цінності не представляють. Так, на жаль, зараз часто «робиться наука», особливо в галузі популяційної генетики.
Антропологічні дані і датування
В останні роки склалася цікава ситуація, коли багато антропологи висловлюють сумнів в «африканське походження людини», але кивають на генетиків, що ми, мовляв, відповіді не маємо, антропологічні дані суперечливі, але от генетики стверджують, що точно знають, що з Африки, тому як ми можемо сперечатися? А ті генетики, які розуміють, що генетичні дані побудовані на піску, точніше, на занадто вільних (фантазійних) інтерпретаціях, кивають на антропологів, що, мовляв, ми розуміємо, що генетичні дані слабкі і часто просто невірні, але от антропологи стверджують, що з Африки, і датування їх на це ж вказують, тому як ми можемо сперечатися? Значить, і у нас все вірно.
Подивимося на твердження, що анатомічно сучасна людина (АСЧ) нібито точно пішов з Африки, і було це близько чи то 40-50 тисяч років тому, чи то 100, чи то 150, чи то 200 тисяч років тому. Нагадаємо, що АСЧ - це той, хто не має помітних архаїчних антропологічних особливостей. Спочатку накидаємо короткий опис обстановки, потім продемонструємо це на конкретному матеріалі. Якщо дійсно коротко, то маємо п'ять основних положень:
(1) всі африканські знахідки стародавніх кісток до приблизно 36 тисяч років тому показують помітні архаїчні особливості;
(2) найчастіше древні кістки настільки фрагментарні, що по них навіть мінімальну антропологічну картину відтворити просто не можна;
(3) часто антропологічна картина кісток в Африці і за межами Африки дуже схожа, і тому не можна сказати, це був вихід з Африки або вхід в Африку;
(4) часто кісткових залишків взагалі немає, і твердження про «анатомічно сучасних людей» робляться на підставі стоянок і кам'яних знарядь, хоча їх цілком могли залишити архантропи, тобто архаїчні люди, що не відносяться до виду «людина розумна»;
(5) датування древніх кісток часто настільки під питанням, що мало хто їх сприймає в буквальному сенсі або навіть просто серйозно.
Почнемо з останнього. На жаль, радіовуглецеві датування на часах більше приблизно 40 тисяч років тому вже майже не працюють, і недавній рекорд з датування - 60 тисяч років тому. Причина проста - період напіврозпаду радіоактивного ізотопу 14С становить 5730 років, тобто 40 тисяч років - це сім періодів напіврозпаду, а 60 тисяч років - це більше десяти періодів напіврозпаду. Метод заснований на вимірюванні співвідношення в біологічних зразках змісту стабільного ізотопу 12С (і трохи 13С, майже в сто разів менше в порівнянні з вмістом 12С) і радіоактивного 14С (при його вихідному вмісті в кількості однієї десятимільярдної відсотка), яке з часом падає, з тим самим періодом напіврозпаду. За 60 тисяч років його вміст зменшується від вихідних 10-10% в 210 рази, тобто ще в 1024 рази. Сучасні прилади такі рівні радіації вже не ловлять, у всякому випадку прилади, які знаходяться в користуванні археологів. Це - приблизно 1 клацання на годину на грам тестованого вуглецю. Звичайний фон значно вище.
Це відкриває «широкі можливості» для помилок, і не тільки помилок. Згадаймо гучну (у вузьких колах) історію з розжалуванням і звільненням зі скандалом німецького антрополога Рейнера Протча (Protsch) за систематичні, як виявилося, фальсифікації датувань стародавніх (і зовсім не древніх) кісток. Після перевірки датувань Протча в 36000 років тому виявилося 7500 років тому, датування 21300 років тому опинилася 2300 років тому, а скелет, датований їм 29400 років тому, виявився залишками людини, яка померла в 1750 році, за 255 років до вимірювань (газета Гардіан). Про це ж писав і відомий міжнародний журнал «Археологія».
Подібних історій чимало в археології, але якщо і відкинути явних фальсифікаторів, яких, звичайно, тільки одиниці, то ймовірність помилок у будь-якому випадку велика. Особливо коли удревнити дуже хочеться, і тим самим увійти в історію археології, а то і культури людства в цілому. Треба сказати, що особливо стародавні біологічні знахідки датуються вже іншими методами, як, наприклад, аргоновим, по відношенню змісту 40Ar/39Ar.
В цілому ж, немає ніяких антропологічних або археологічних доказів «походження сучасної людини в Африці», як і немає ніяких доказів, що кам'яні «інструменти» і «індустрії», знайдені в Європі чи в Євразії в цілому, були привнесені з території на південь від Сахари. Всі заяви про виявлення кісткових залишків «анатомічно сучасної людини» давністю більше 50 тисяч років, і тим більше більш 150 тисяч років, і тим більше на південь від Сахари, є просто спотвореними або невірними з самого початку. Досить повний огляд на цю тему буде незабаром опубліковано відомим австралійським антропологом Робертом Беднаріком (у пресі, Advances in Anthropology).
Про відсутність таких знахідок на південь від Сахари свідчать ряд робіт, наприклад (Grine et al, 2007; Grine et al, 2010). Всі відомі знахідки кісток такої давнину носять явні архаїчні особливості, починаючи з Omo Kibish 1 (195 тисяч років тому, Ефіопія, знайдені фрагменти черепних кісток, лицьових кісток мало), Omo-2 (показаний ряд примітивних, архаїчних особливостей), Herto (154-160 тис років тому, дуже архаїчна кісткова будова, надто відрізняється від АСЧ), загалом, всі скелетні залишки, що датуються 100-200 тисяч років тому, і фактично раніше 35 тисяч років тому, є архаїчними (Rightmire, 2009). Багато які взагалі не мають збережених лицьових кісток. Навіть череп Hofmeyr з Південної Африки, що датується 36 тисяч років тому, має архаїчні особливості (Grine et al, 2007; Rightmire, 2009; Tattersall, 2009).
Американський антрополог Райтмайр повідомив - «ні копалини Herto, ні інші з пізнього Плейстоцену, як Klasies River в Південній Африці чи Skhul/Qafzeh в Ізраїлі, не мають аналогій із сучасними популяціями. Їх черепа являються робастними, і лише починаючи з ~ 35 000 років тому починають з'являтися люди сучасної анатомічної морфології» (Rightmire, 2009). Він вважає, що «анатомічно сучасні люди» еволюціонували в Африці, хоча цей процес «погано зрозумілий». Йому вторить Майкл Хаммер (Hammer et al, 2011) - «копалини гомінин, що показують комбінацію архаїчних і більш сучасних особливостей, постійно зустрічаються в Африці на південь від Сахари і на Близькому Сході аж до приблизно 35 тисяч років тому». Таким чином, постійні посилання прихильників концепції «з Африки», що в Африці виявлені кісткові залишки анатомічно сучасних людей з датуваннями між 160 і 200 тисяч років тому, є помилковими. Маніпуляції і спотворення тривають і тут.
Майкл Хаммер, в недавньому минулому активний захисник концепції «з Африки», став висловлювати сумніви у виході «з Африки», але став це виражати вже в 2013 році, коли стало ясно, що ситуація з «виходом з Африки» заплуталася донезмоги. Хаммер закінчує свою статтю в журналі «Сайнтифик Амерікен» (травень 2013 року) наступними словами: «Багато вузлів залишилися нераспутаннами. Але одне ясно - коріння сучасної людини йдуть не тільки до єдиної пращурної популяції в Африці, але до популяцій Старого Світу» (тобто Європи чи Євразії - АК).
І це видається цілком розумним. Вивчаючи історію стародавнього світу, ми постійно зустрічаємося з мобільними міграціями, в тому числі й досить далекими міграціями. Якщо за версією прихильників «виходу з Африки» людина всього за 10 тисяч років досягла Австралії, то важко собі уявити, що він сиднем сидів 200 тисяч років і не розійшовся по всьому світу, в тому числі і до Африки і з Африки, причому багато разів. Як вийшло, що концепція одностороннього «виходу з Африки» так агресивно нав'язувалася суспільству і так швидко їм опанувала, має викликати тривогу - і щодо агресивного, безапеляційного нав'язування, і щодо настільки безвольного сприйняття громадськістю. І адже що особливо повинно турбувати - адже ніяких надійних підстав у «виходу з Африки» не було і немає.
Резюмуючи, повторимо ще раз - наші пращури з Африки в останні 200 тисяч років не виходили. Інакше кажучи, ті , які з Африки виходили, не породили сучасне неафриканське людство. Це показує весь комплекс отриманих даних - і генетика, і антропологія, і археологія, і ДНК-генеалогія. Насправді і не тільки в останні 200 тисяч років, а й раніше. Вивчення ДНК викопних кісток нендертальців показало наявність в них рецептора меланокортин (MCR1), причому у варіанті, який задає світлу шкіру і руде волосся (Lalueza - Fox et al, 2007). Автори вважають, що неандертальці за кольором волосся приблизно відповідали сучасним європейцям - від темноволосих до блондинів. До того ж не виявлено ніяких показників, що неандертальці були негроїдами. Дійсно, в Африці слідів неандертальців не виявлено. А оскільки наші найближчі пращури з неандертальцями були загальні, оскільки неандерталець - наш племінник, то і «батько» неандертальця і «брат» нашого «батька» теж, швидше за все, мав світлу шкіру і жив не в Африці. Було це десь в інтервалі 300-600 тисяч років тому. Залишається, втім, нерозгаданим, як наші светлошкірі брати, з якими ми розійшлися приблизно 160 тисяч років тому, вижили, потрапивши в Африку, і як придбали темний колір шкіри, але відповідь знайдеться теж в області генетики, у регуляції біосинтезу меланіну. Але це вже інша історія.
література
Анатолій А. Кльосов,
професор, доктор хімічних наук
Пізнаємо природу людини. Якщо прагнеш щось пізнати - поглянь, звідки воно виникло.